大腸で水分が吸収されることで糞便が固くなります。大腸の後ろのほうが水分が吸収される割合が大きいので固くなるわけですね。教科書的な知識としてはそうですが、実際それを実感するのは、犬の散歩で犬に糞をさせるときです。1回の散歩で2回くらい糞をしますが、1回目が固くて2回目が柔らかいです。教科書の知識と実体験が結びつく瞬間でした。
「未分類」カテゴリーアーカイブ
ミュラー管の発生
ミュラー管の発生の発生はまだその全貌が明らかにされてはいないようです。しかし、
Cellular mechanisms of Müllerian duct formation in the mouse Grant D. Orvis, Richard R. Behringer Developmental Biology Volume 306, Issue 2, 15 June 2007, Pages 493-504 Developmental Biology https://www.sciencedirect.com/science/article/pii/S001216060700228X?via%3Dihub
に詳し解析があります。
- In the first phase, cells of the coelomic epithelium are specified to a Müllerian duct fate through an unknown mechanism. This specification can be identified by the expression of Lim1 in cells of the coelomic epithelium (Kobayashi et al., 2004).
- After specification, Wnt4 expression from the mesonephros or coelomic epithelium, induces the invagination of these specified cells (Kobayashi et al., 2004, Vainio et al., 1999).
- when the Wolffian duct is disrupted at a specific point in ovo, the Müllerian duct is unable to grow past that point (Gruenwald, 1937).
https://www.sciencedirect.com/science/article/pii/S001216060700228X?via%3Dihub
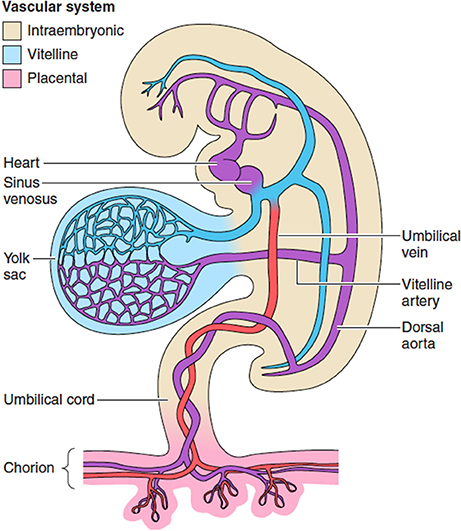
臍帯 umbllical cordに含まれるもの?卵黄嚢はどこに?
臍からおしっこが漏れ出すことがあるというのを教科書で読んで、臍と膀胱がつながっているのかと感銘を受けたのですが、あれ?卵黄嚢も臍帯に含まれていなかったっけ?と疑問が生じました。もしそうなら腸管と臍が繋がるんじゃないかという疑問が湧いたわけです。
調べてみたら、そういうことはありませんでした。
発生初期の胚の図を見ると卵黄嚢も臍帯に含まれているような描き方がされていると思います。しかしそれは早期だけのようです。”primitive umbilical cord”と、”definitive umbilical cord”というように言葉も使い分けられていました。”primitive umbilical cord”は umbilical ringとも言われるようです。
primitive umbilical cord
下のラングマンの教科書からの図では、腸管の伸長に伴う生理的なヘルニアが生じている時期を示しているので、断面図で腸管が描かれています(これは一時的なもの)。
Primitive umbilical cord and contents. A. A 5-week embryo showing structures passing through the primitive umbilical ring. B. The primitive umbilical cord of a 10-week embryo. C. Transverse section through the structures at the level of the umbilical ring. D. Transverse section through the primitive umbilical cord showing intestinal loops protruding in the cord. (Reprinted with permission from Sadler TW. Langman’s Medical Embryology. 13th ed. Philadelphia, PA: Wolters Kluwer Health; 2015.) https://renaissance.stonybrookmedicine.edu/system/files/Umbilical%20Cord%20Disorders.pdf

definitive umbilical cord もしくは単にumbilical cord 臍帯
Diagrams showing the (A) folding of the embryo and formation of the primitive umbilical cord and (B) definitive umbilical cord.
https://www.researchgate.net/publication/305720901_Anatomy_and_embryology_of_umbilicus_in_newborns_a_review_and_clinical_correlations




上の図では発生の早期と後期を示していて、一目瞭然でわかりやすいと思います。
https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2018.00373/full
https://www.researchgate.net/publication/363584025_Bovine_omphalocele_errors_in_embryonic_development_veterinarian_importance_and_the_way_forward
https://www.researchgate.net/publication/301814976_Abdominal_Wall_and_Umbilicus-_Essentials_of_surgery-_Sunil_Chumber



Allantoi 尿膜の発生のその後
以下、ChatGPT4oによるまとめです。
The allantois is functionally obliterated early in development but leaves remnants that persist after birth. Here’s how it works:
Timeline of Allantois Obliteration 消滅:
- Early Embryogenesis:
- The allantois emerges as a small endodermal outpouching of the hindgut.
- It extends into the connecting stalk (which contributes to the formation of the umbilical cord) and plays a role in early gas exchange and the development of the umbilical arteries and veins.
- By the end of the first trimester, its role in development diminishes as the placenta takes over waste exchange.
- Obliteration during Development:
- The lumen of the allantois is obliterated during the second trimester, and it becomes a fibrous cord known as the urachus(尿膜管), which runs from the bladder to the umbilicus.
- By birth, the allantois is no longer a functional structure, but its remnant persists as the median umbilical ligament (正中臍索)in postnatal life.
- Postnatal Remnants:
- The median umbilical ligament extends from the apex of the bladder to the umbilicus and is the fibrous remnant of the allantois.
- If the obliteration of the allantois is incomplete, it can result in abnormalities like a urachal cyst, urachal sinus, or even a urachal fistula, which might require surgical correction.
腎臓の働き、構造、位置など
腎臓はどこにある?
- 腎臓は第12胸椎から第3腰椎の間にあります。
- 右の腎臓は、上部に肝臓があるために左の腎臓より少し低い位置にあります。
- 腎臓は、背骨と第12肋骨(一番下の肋骨)に囲まれた背中側に在ります。
- 腎臓は左右にあり、腎臓の上下の長さはだいたい10-12cm程度、一つの重さは120g程度。
https://www.tamana-uro.com/2009/04/16/%E9%99%A2%E9%95%B7%E3%81%93%E3%82%89%E3%82%80%E3%80%80%EF%BC%9A%E3%80%80%E8%85%8E%E8%87%93%E3%81%AE%E3%81%8A%E8%A9%B1%E3%80%80-%E3%81%9D%E3%81%AE1/
性同一性障害は性ホルモンの脳内作用によってどこまで説明がつくのか
総説
- Brain Sex Differences Related to Gender Identity Development: Genes or Hormones? Int J Mol Sci. 2020 Mar 19;21(6):2123. doi: 10.3390/ijms21062123
脳の性分化のメカニズム:アンドロゲンの作用
性同一性障害の典型例
- 胎児期に男性ホルモン(テストステロン)が多いと脳が男性化し出生後も男性としての性行動(少ないと女性化)
- 染色体・性腺・性器が男性でも、胎児期に男性ホルモンが少ないと女性としての性行動
- 染色体・性腺・性器が女性でも、胎児期に男性ホルモン(テストステロン)の量が多いと、男性としての性行動
https://caran-coron.jp/column/128/
ホルモン療法
男性が、女性ホルモンの補充療法を受けた場合、下のリンクの説明によれば、乳房が大きくなり、乳腺と脂肪が増加しし、体脂肪の付き方が変わって女性らしい顔つきとともに腰回りの脂肪が増え、勃起力が低下し、性欲も減少し、精子量は減少するのだそうです。また外見だけでなく、性格が女性的に変化し、涙もろくなどなるそうです。その他、男性型脱毛症が改善して、髪の毛が細くなり密度が高まるなど頭髪が女性らしく変化するのだそう。成人男性の場合に、声や骨格まで女性化することはないそう。
女性が男性ホルモンの補充療法を受けた場合は、乳腺の収縮と皮下脂肪の減少によって乳房が縮小するほか、皮下脂肪が減少し、筋肉が増加し、内臓脂肪が増加するそうです。また、生理が止まります。外見だけでなく、内面的な変化として、性格が男性的になるのだそうです。そのほか、ひげが発生し、頭髪が減少します。成人女性の場合に、骨格の変化はあまり起きないそうです。
性ホルモン注射療法は、FTM(Female-to-Male)の場合にはアンドロゲン製剤を、MTF(Male-to-Female)の場合にはエストロゲン製剤を注射 https://www.azabujuban-clinic.jp/gender_dysphoria/
語句
トランスセクシャル:身体的性と性自認が一致せず、医学的な処置(性別適合手術やホルモン注射など)によって、身体と自認する性別の一致を望む人々のこと、もしくはそのような処置をすでに受けた人たちのこと。
- https://www.ikecen.com/gid
アンドロゲンから変換されてできるエストロゲンの男性的な行動に対する影響
男性ホルモン(アンドロゲン)が男性的な行動をつくりだすということなら話は単純ですが、実際には、アンドロゲンであるテストステロンは細胞内のアロマターゼによってエストロゲンに変換されて、エストロゲンの作用を持ちます。ですから、単純にテストステロン=男性ホルモンと考えて実験結果を解釈するわけにもいかなそうです。
- The Androgen Receptor Governs the Execution, but Not Programming, of Male Sexual and Territorial Behaviors Scott A. Juntti 1 6 , Jessica Tollkuhn 3 6 , Melody V. Wu 1 , Eleanor J. Fraser 2 , Taylor Soderborg 3 7 , Stella Tan 3 8 , Shin-Ichiro Honda 5 , Nobuhiro Harada 5 , Nirao M. Shah Neuron Volume 66, Issue 2, 29 April 2010, Pages 260-272 https://www.sciencedirect.com/science/article/pii/S0896627310001959?via%3Dihub
- Estrogen Masculinizes Neural Pathways and Sex-Specific Behaviors Cell Volume 139, Issue 1, 2 October 2009, Pages 61-72 Melody V. Wu 1 , Devanand S. Manoli 4 , Eleanor J. Fraser 2 , Jennifer K. Coats 1 , Jessica Tollkuhn 3 , Shin-Ichiro Honda 6 , Nobuhiro Harada 6 , Nirao M. Shah https://www.sciencedirect.com/science/article/pii/S0092867409009167?via%3Dihub
性ホルモン(アンドロゲン、エストロゲン)とストレス反応との関連性
- Roles for androgens in mediating the sex differences of neuroendocrine and behavioral stress responses Biology of Sex Differences volume 11, Article number: 44 (2020) 総説 https://bsd.biomedcentral.com/articles/10.1186/s13293-020-00319-2
生殖器の発生発達:男性と女性との中間の表現型、遺伝型との不一致などについて
未分化な性腺における遺伝子発現
Gonadal primordia is observed in humans in week 5 of gestation, being under the control of WT1, NR5A1, NR0B1, CBX1/2, LHX9, EMX2, GATA4, and SIX1/4 [15].
Studies in mice have shown that, at this moment, genes that are associated with Sertoli (testicular) (SOX9, FGF9, PGD2) or granulosa (ovarian) (WNT4, RSPO1, CTNNB1, FST) differentiation are expressed at similar levels in both the XY and XX fetus [16,17,18].
https://www.mdpi.com/2075-4418/11/8/1379
さりげないモニタリング unobtrusiv sensingによる健康の維持増進
自分は父を大腸癌で亡くしましたが、もし大腸癌検診(大腸カメラ)などを定期的にしていれば外科的に取れる段階で見つけられたのではないかと思います。病院嫌いの父でしたので、早期検診などという概念とは無縁の生活をしていました。それまで健康に暮らしていたのに、ある時突然ひどい便秘になってしまい検査した結果、大腸癌が大きくなって大腸を塞いでいたために便が出てこれなくなっていたのでした。即入院してステントを入れて便通はできるようになりましたが、がんに関してはすでに肝臓や他の部位にまで転移していて、なすすべがない状態になってしまっていました。
職場の定期健診ではオプションで検便があります。潜血がないかどうかを調べて、もしあった場合には大腸癌が疑われるので大腸カメラをやったほうがいいというものです。尿検査で採尿するだけでもうっとおしいと感じているのに、自分のうんちを採取するなんてそんな面倒なことはやりたくないと、何年もの間検便は避けてきたのですが、父を大腸癌で亡くし、癌が家族性(リスクファクター)であることを考えると、あるとき突然死の宣告を受けるのも厳しいなと思って、今度から検便のオプションを選ぶようにしています。
しかしトイレで便を採取するのは面倒です。いっそトイレに潜血を自動的に検査するシステムが組み込まれていればよいのにと思いました。遠くない将来、そのようなスマートトイレが市場に出てくるのではないかと思いますが、現状はどうなっているのでしょうか。検便もそうですが、それ以外にも、健康管理を自動測定して警告を発してくれるシステムがあればいいと思います。家に防犯カメラなどのセキュリティをつける人がいますが、同様に常時自分や家族の健康状態が(精神面も含めて)測定できれば、なにかと便利そうです。もちろん個人情報を記録することになるので、その管理という点でいろいろクリアすべき問題はあるでしょう。
能動的に検査を受けるのは面倒ですので、普段の生活を邪魔されずに勝手に健康状態をモニタしてもらえると有難いと思います。そんな考え方が実はすでに世の中にあることを知りました。
「さりげないセンシングと日常人間ドックで実現する自助と共助の社会創生拠点」という東北大学および共同研究する企業からなる研究グループのプロジェクトです。科学技術振興機構(JST)の助成事業センター・オブ・イノベーション(COI)プログラムで行われたプロジェクトだったようです。
- JSTトップ > 【終了】COIプログラムトップ COIプログラムは令和4年3月末をもって終了いたしました。(事業実施期間:平成25年~令和3年)
-
さりげないセンシングと日常人間ドックで実現する自助と共助の社会創生拠点 東北大学

さりげない尿の成分測定
尿中のナトリウムイオンとカリウムイオンの濃度の比(ナトカリ)が血圧と相関するのだそうです。Na/K の比が高いほど高血圧である可能性が高いのだそう。よってナトカリをモニタすることで、高血圧ぎみですよと注意を促してくれるわけです。
2024 年 10 月 8 日 日本高血圧学会 尿ナトリウム/カリウム比(尿ナトカリ比)ワーキンググループ コンセンサスステートメントの発表について 日本高血圧学会は、日本人のための尿ナトカリ比の目標値と適切な評価方法を提唱するた め、機関誌 Hypertension Research(JSH STATEMENT)にて 2024 年 10 月 8 日(火)にコン センサスステートメントを公表いたしました。 https://www.jpnsh.jp/data/pressrelease_241008.pdf
- センター・オブ・イノベーション(COI)プログラム 終了報告書 研究開発期間:平成 25 年度~令和 3 年度 拠 点 名: さりげないセンシングと日常人間ドックで実現する 自助と共助の社会創生拠点 中核機関: 東北大学(72ページPDF)https://www.jst.go.jp/coi/hyoka/data/110_tohoku_hokoku.pdf
- 尿ナトカリ比と高血圧リスクの関連 https://healthcare.kagome.co.jp/column/natokari
ピロリ菌と胃がんとの関係
大分大学研究紹介:医学部医学科 解剖学講座 教授 濱田 文彦 大分大学研究マネジメント機構 産学官連携推進センター チャンネル登録者数 194人
研究マネジメントとは
最近、大学における研究マネジメントの重要性が謳われるようになってきていますが、そもそも研究マネジメントとは何でしょうか?研究マネジメント人材に求められる仕事内容はどのようなものなのでしょうか。
民間企業における研究開発マネジメントは商品化が最終目標だと思いますが、大学における研究マネジメントはゴールが多少異なると思います。最終目標(問われる成果)は、論文数、企業との共同研究実施実績などのようです。
本事業は、大学等が、研究経営システムを強化するため、リサーチ・アドミニストレーター(以下、「URA」という。)及び産学官連携コーディネーター(以下、「CD」という。)等に対して、研究経営能力等の育成に向けた研修の実施により、産学官連携活動の持続可能な体制整備及びURA、CD等を含む研究支援業務を担当する人材の資質向上を図るとともに、研究マネジメント人材育成のための体制整備及び強化を行うために必要となる要素及び課題を抽出し、調査・分析を行ったものである。その結果を以下の通り取りまとめた。(研究マネジメント人材の育成に向けた調査分析事業 文部科学省)
研究開発マネジメント人材とは、JSTのファンディング事業に特化し、研究成果最大化に向け、事業における研究領域等の推進業務を行う人材です。採用された方にはイノベーションの触媒、目利きとして、研究開発プログラムの公募選考・管理・成果展開等に携わっていただきます。本制度のもと、将来的には、研究者のパートナーとなって様々な関係者をつなぎ、研究活動全体をマネジメントするプロデューサー的人材(研究開発マネージャー)へと育っていくことを目指します。事例1「青色LED」JST職員が早期に企業連携を説得 日本での実用化に成功1985年、研究の意義をいち早く見出す「研究に集中し他に時間を取られたくない」赤﨑教授を訪問 産学官連携によるGaN青色発行ダイオード開発の必要性を粘り強く説く これをきっかけに、豊田合成との産学連携プロジェクトが立ち上がり、後にノーベル賞を受賞する青色LEDの実用化にむけた取り組みが開始。事例2「iPS細胞」 京都大学教授 山中伸弥 JST職員がALL JAPAN研究体制構築に動く 山中教授は研究に専念、国際社会をリード ヒトiPS細胞樹立後、多忙を極めた山中教授 機を逃さずJSTが研究をマネジメントできる職員を長期派遣 右腕としてセンター立ち上げに貢献、滞っていた研究支援を担当。 (研究費獲得、ルール策定・学外連携、論文相談等) iPS細胞研究所(CiRA)として世界に先駆け臨床応用へ発展。(研究開発マネジメント人材の趣旨 JST)
本報告書において、「研究開発マネジメント」とは、大学や研究機関において、科学技 術イノベーションの創出に向けて行われる、スキル標準に示された研究戦略推進支援業務 6、プレアワード業務、ポストアワード業務、教育プロジェクト支援等の関連業務を中心 として、近年求められる、研究セキュリティ/インテグリティ、倫理的・法制度的・社会 的課題 ELSI(ethical, legal and social implications/issues)、スタートアップ支援、ファ ンドレイズ、オープンアクセス/オープンサイエンスへの対応等も含む概念とする。 研究開発マネジメントが充実することにより、研究はもちろんのこと、産学官連携、教 育、組織運営等の機能を最大限、効果的に果たすことに寄与することが期待される。
科学技術イノベーションの創出に向けた研究開発マネジメント業務・人材に係る課題の整理と今後の在り方 資料2-2 科学技術・学術審議会 人材委員会(第 102回) 令和6年6月24日(39ページPDF)
大学における研究マネジメントの事例集
- 研究マネジメントの現状と課題(PDF) 2023年 東工大、筑波大、立命館大、中央大の事例紹介
自分なりにですが簡単にまとめてしまえば、研究マネジメントとは、大学としてのビジョン(大学の社会的使命)にのっとって制度(支援組織の整備)を整えて目標を設定し、お金(外部・内部の研究費)と人(研究者およびそのほかの支援者など)をやりくりして、モノ(既にある研究設備など)と情報(誰がどんな研究成果を上げているのか、どこにビジネスチャンスがあるのか、どんな企業が何をやりたがっているのか)も活用して結果を出す(論文、資金獲得、科学研究の成果の社会実装)ということのようです。
研究評価 IR
研究者個人の評価指標の確立:科研費獲得件数、共同研究の実施件数、トップ10%論文数、ハイインパクトジャーナル掲載数
研究支援制度の充実
科研費不採択者に対して審査結果に応じて助成
産学連携の推進
研究シーズ集の作成、公開、キーワード検索システムの実装
研究マネジメントの成果の指標
論文数
トップ10%論文数
国際共著論文
外部資金獲得:公的研究費(科研費ほか)
大型予算 特別推進研究、基盤研究(A)
さきがけ
申請件数、採択件数、採択順位
企業との共同研究
組織対組織(大学と企業)の共同研究の件数
産学連携研究による研究費収入 受託研究費、共同研究費
受賞
参考
- Research Development at MIT https://research.mit.edu/research-development
- Research management is a broad field involving the coordination of numerous processes to move research efforts forward, from setting goals and obtaining approval to monitoring progress and ensuring compliance. For research administrators, it is vital to understand what this entails so they can manage projects effectively and make better decisions. https://cayuse.com/blog/5-key-areas-of-research-management/
排便の仕組みと科学
肛門の解剖学
肛門の周りには内肛門括約筋がありさらにそのまわりには外肛門括約筋が存在しています。直腸の圧がたかまると伸展刺激が副交感神経系の骨盤内臓神経に伝わって、それが脊髄の神経細胞を経由して内肛門括約筋を弛緩させます。一方、外肛門括約筋は運動神経である陰部神経の支配を受けており陰部神経が外肛門括約筋の弛緩や収縮を制御します。陰部神経の細胞体は脊髄にありますが、その細胞に対して脳から排便命令を伝える神経終末がシナプスを作って、情報を送ります。
便意が発生するしくみ
直腸内圧が増加するとその刺激が骨盤神経により仙髄の排便中枢につたえられ、脊髄反射による遠心性線維を介して直腸の蠕動運動が高められます。そして、内肛門括約筋を抑制して弛緩させることで、糞便の排出をうながします。直腸内圧の増大による刺激は、また大脳にも送られて便意をもよおします。
排便を我慢する仕組みと排便の仕組み
便意が生じたからといっていつでもどこでも排便していいわけではないので、ここで人間は意思の力をもって、排便をコントロールするわけです。意図的にもしくは反射的に、外肛門筋を収縮させることにより、排便をこらえます。トイレに言って便座に腰かけて(あるいは和式トイレなら便器にまたいで)、排便してOKな状況になれば、「いきみ」(Pushing)などによってあるいは陰部神経を介して外肛門括約筋を弛緩させて排便に至ります。陰部神経ー外肛門括約筋は、随意運動が可能な骨格筋と同様に「運動神経ー横紋筋」のシステムであり意思によるコントロールができるというわけです。
正しい排便の方法
For a perfect poo
排便時の姿勢 To help open your anorectal passage
- knees higher than your hips
- Lean forward
- Bulge out your tummy while keeping a straight back
排便時の動作
- relax your back passage(おしりの出口をリラックスさせ) and let go
- Don’t hold your breath or strain(りきむ).
https://www.abc.net.au/news/2019-07-18/reasons-we-strain-to-poo-and-what-to-do-about-it/11316776
トイレットペーパー不要の完全無欠な排便に関する議論
After I defecate and attempt to wipe, there’s never anything on the toilet paper.
It’s called ghost poop. Your poop is so well made and compacted into the most anal-friendly poop rocket, that it has no time to bother your little bum hole.
https://www.quora.com/After-I-defecate-and-attempt-to-wipe-theres-never-anything-on-the-toilet-paper-This-has-been-going-on-for-weeks-now-Is-this-normal
参考
- しくみが見える体の図鑑 2012年 株式会社エクスナレッジ 代表取締役社長:三輪浩之 https://www.xknowledge.co.jp/book/9784767814421 この本は日本語の名称だけでなく英語の名称が付記されていて、英語圏の情報にアクセスする際に役立ちます。また日本語の解剖学用語は読み方が難しいものが多いのですが、この本ではふりがなが振られており、それもまた非常に役立ちます。この図鑑は臓器ひとつひとつの「見せ方」が上手くて、理解しやすいなあと感心して出版社を見てみたら、エクスナレッジという自分が知らない会社でした。ネットであれこれ見てみたら、「わかりやすさ」へのこだわりがあればこそなのかと納得。
- 雑誌『建築知識』は1959(昭和34)年に発刊。今年で創刊から60周年を迎える「読者の9割近くは1級建築士等の建築専門家」という、コテコテの建築専門誌であります。発行元のエクスナレッジは、建築だけにとどまらず、デザイン・生物・コンピューター・ビジネス関連など、多岐にわたるジャンルで書籍を刊行し、生物ジャンルでは、猫以外にもサルトリイヌペンギンクモイカタコウサギの書籍も数多く手がけています。特に、猫書籍のクオリティは高く、書店営業の折には「猫の出版社でしょ?」と言われることもあるほど。https://home.kingsoft.jp/news/amusing/nekojournal/11029.html
- 『日本の最も美しい教会』 https://www.christiantoday.co.jp/articles/23298/20170223/the-most-beautiful-church-in-japan.htm
- 2021.08.08 12:00 『建築知識』編集長に訊く、バズる専門誌の作り方 大きな転換点となった“猫のための家づくり” 休日に30キロくらい歩いています。その時間って、何も考えてないようでいていろいろ考えますし、自然や町並みを見て何かしらヒントを得ていたりもします。そうした意味で、「無駄な時間」って編集者にとってはすごく大事なものだと思うのです。https://realsound.jp/book/2021/08/post-829818_2.html
- 建築雑誌『建築知識』、建築基準法を「萌えキャラ」に擬人化 インタビュー エトセトラ 長井 雄一朗 公開日:2017.12.18 堅い雑誌というイメージが強い同誌だが、今年の12月号では、建築基準法などの建築法規を「萌え絵キャラクター」で表現し、物議をかもしている。 https://sekokan-navi.jp/magazine/12023
- 2017年8月号 広報会議 専門メディアの現場から 記事シェア 「猫」特集で売り切れ店続出 建築家100人以上に取材する実務誌 エクスナレッジ『建築知識』 『建築知識』は建築業関連従事者に専門知識を伝える実務誌だ。しかし本年1月号では盛り上がる猫ブームを捉え、「猫のための家づくり」を特集したところ売り切れ店が続出。ネットでプレミアがつくほどの人気を呼んだ。「対象はあくまで建築家でしたが予想を超える反響。猫を飼う一般女性も多く購入されたようです」と編集長の三輪浩之氏は振り返る。https://mag.sendenkaigi.com/kouhou/201708/scene-of-media/011205.php
- 2021.08.08 12:00 『建築知識』編集長に訊く、バズる専門誌の作り方 大きな転換点となった“猫のための家づくり” 文・写真・取材=木下恵修、画像提供=エクスナレッジ 『解剖図鑑』シリーズはこれまでに40冊以上出版していますが、その多くを担当しました。『日本の神様 解剖図鑑』や『日本の戦争 解剖図鑑』『百人一首 解剖図鑑』など、建築とは関わりのないタイトルがいくつもありますが、実はいずれも『建築知識』と同じつくり方をしています。https://realsound.jp/book/2021/08/post-829818.html
- 『建築知識』ヒット企画は出版業界仰天の「働き方改革」が源⁉編集部2021年07月15日 編集の根底に流れるのは、「実務に必ず役立つテーマを扱う」というスタンスである。その際に表現方法を工夫すれば、一般読者も取り込めるのでは……と編集しているそうだ。https://kensetsu-hr.resocia.jp/article/interview_kenchikuchisiki
- 足が速くなる解剖図鑑 2024/10/3 高林 孝光 (著) エクスナレッジ (2024/10/3)