細胞の極性 cell polarity に関わる分子は、結果的に個体の体軸を決めたり様々な役割を担います。細胞極性を生み出す分子メカニズムは、生命現象の根源的な理解に欠かせません。
Geraldine Seydoux (Johns Hopkins / HHMI) 1: From Egg to Worm: How to Create a Body Axis Science Communication Lab チャンネル登録者数 18万人 11:39から遺伝学的スクリーニングによって細胞の非対称性に関わる分子を同定する話になります。
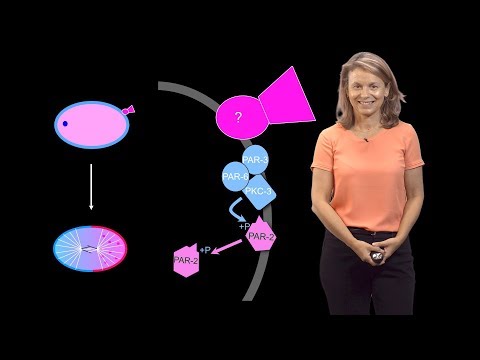
多くの重要な分子はC. elegansの突然変異体スクリーニングで同定されました。それらはpar-1, par-2, par-3, par-4などと名付けられています。parはpartitioning-defectiveの意味。 非対称に分裂するはずのところで、対称に分裂する表現型をもつ変異体として同定されました。そのため、名称がおなじparでも番号がことなる分子は相互に無関係です。すなわち、相同性があるとか、ファミリーを構成するわけではありません。
まだ1細胞の時期に将来の前側に局在する分子としては、PAR-6, PAR-3, PKC-3 (atypical PKC; aPKC)があります。後方側に局在する分子としてはPAR-1, PAR-2があります(上の動画13:35)。
- 前側に局在する分子: PAR-6, PAR-3(lipid binding), aPKC(kinase)
- 後方側に局在する分子: PAR-1(kinase; lipid binding), PAR-2(lipid binding)
これらの分子の特徴として、PAR-3とPAR-1、PAR-2は脂質結合ドメインを持ちます。つまり細胞膜上に局在できます。また、PKC-3とPAR-1はキナーゼドメインを持ちます。つまりリン酸化酵素としての働きがあります(動画15:26)。
前方のPAR群と後方のPAR群は競合的に働きます。その仕組みは見事なもので、キナーゼが相手の群れの分子の脂質結合ドメインをリン酸化して脂質と結合できなくするというものです。前方群のPKC-3(キナーゼ)は後方群に属するPAR-1とPAR-2の脂質結合部位をリン酸化します。逆に、後方群のPAR-1(キナーゼ)は、前方群に属するPAR-3の脂質結合部位をリン酸化します(動画16:57)。
受精前の線虫の卵では「前方群」の分子(PAR-3, PAR-6, PKC-3)が細胞全体に存在していて局在はしていません。PAR-2の脂質結合部位をリン酸化した状態にたもっているからです。
受精により精子が侵入することで、この状況がどう変化するのでしょうか。精子は中心体を持込み、中心体は微小管を形成する中心として働くので微小管が発達し、PAR-2は微小管と結合することによりリン酸化部位が「保護」されて、細胞膜に結合できるようになります。PAR-2は相棒のPAR-1(キナーゼ)をリクルートし、PAR-1がPAR-3をリン酸化します。こうして精子が侵入したあたりの膜は「後方化」するわけです。
後方化する部分は細胞の半分近くの広さまで広がりますが、それにはまたひとつ巧妙なしくみがあるそうで、アクトミオシン系が前方群のPARタンパク質たちを前方へ移動させるのだそうです(動画26:10)。
この動画は線虫の話でした。動物種が異なれば同じ分子の役割も異なる可能性があります。哺乳類などでは、タイトジャンクションにPAR-6, PAR-3, aPKCが複合体として存在するといった報告があるようです。哺乳類の発生を考えるうえで、線虫で得られた知見はそのままは適用できなくても、インスピレーションを与えてくれるでしょう。