Th1/Th2バランスとは
病態の説明などにおいてしばしばTh1/Th2バランスといった言葉が登場します。これはヘルパーT細胞の亜型であるTh1細胞とTh2細胞のそれぞれの働きの拮抗をあらわした概念です。Th1細胞が細胞性免疫を亢進させる働きを持つのに対して、Th2細胞は体液性免疫を亢進させる働きを持ちます。また、Th1細胞はインターフェロンγを分泌してTh2細胞に働きかけその活動を抑制します。逆に、Th2細胞はインターロイキン4やインターロイキン10、TGF-βなどを産生してTh1細胞を抑制するように働きかけます。
- リッピンコット免疫学172~173ページ Th1/Th2パラダイム、253ページ Th1/Th2バランス
- The Th1/Th2 paradigm: still important in pregnancy? 03 May 2007
- Revisiting the Th1/Th2 paradigm Muraille 1998 (PDF)
- The Th1/Th2 paradigm Sergio Romagnani June 1997 Immunology Today
Th1/Th2パラダイムは非常にわかりやすい仮説(パラダイム)なので魅力的ですが、必ずしもこのような単純な図式では説明がつかないこともあるため、免疫学の教科書を見た時に取り扱い方は様々です。リッピンコットの教科書では、「モデル」としてしっかりと紹介されていました。
最近の教科書を見ると、Th1/Th2パラダイムを紹介していないものも多いようです。自分は、科学は事実の羅列ではなく、どう理解するか、概念の確立だと思っているので、かりに訂正が必要であったとしてもこのような明確な概念、パラダイムが研究の進展において果たした役割は説明してほしいと思います。
Th1/Th2パラダイムの趨勢
Th1/Th2細胞のバランスが、生体の免疫応答の性質を決めるというこの仮説はその後数十年間、免疫学の世界を支配するパラダイムになりました。(私的免疫学ことはじめ (2) Th1/Th2パラダイム 2020年7月30日 医局ブログ)
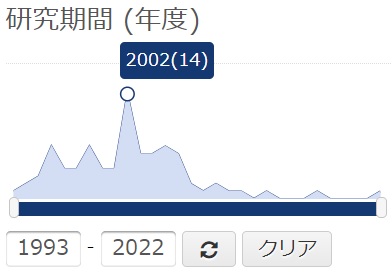
科研費の採択課題をみてみると、Th1/Th2バランスを前面に推した計画は2002年ころがピークだったようです。

-
-
- Th1/Th2バランスをターゲットとしたPDE5阻害薬による流産改善効果の検証 2022-04-01 – 2025-03-31
- 舌下免疫療法におけるセマフォリン4Aを介した Th1/Th2制御の解明 2017-04-01 – 2020-03-31
- 被嚢性腹膜硬化症におけるTヘルパー細胞の役割の解明と新規治療法の開発 2013-04-01 – 2016-03-31
- Thバランスの制御による難治性喘息に対する新規治療法の探索 2011 – 2013
- Th17/Th1/Th2細胞優位発現マウスを用いた免疫複合体腎炎の病態解析 2010 – 2012
- メモリーTh1/Th2細胞の形成と機能維持のエビジェネティック制御に関する研究 2009 – 2011
- Th1 Th2 バランスの制御による魚類のウイルス病に対する細胞性免疫誘導 2009 – 2010
- Th1/Th2バランスの破綻として捉える薬剤性肝障害 2008 – 2010
- Th1/Th2細胞分化におけるケモカインCCL21、CCL19の役割の解析 2007 – 2008
- 乳幼児期の細菌刺激および化学物質曝露による成長後のTh1/Th2バランスへの影響 2007 – 2009
- 抗うつ薬応答性に及ぼすTh1/Th2サイトカイン遺伝子多型の影響 2006
- チロシンキナーゼTxkによるマスト細胞のTh1/Th2応答の制御機構 2006 – 2007
- Tim分子によるTh1/Th2反応制御機構の解明 2006 – 2007
- ナノ粒子(酸化チタンおよび酸化亜鉛)のTh1/Th2/Th3免疫応答に与える影響 2006 – 2007
- 魚類のTh1/Th2バランス制御機構の解明 2006 – 2008
- セマフォリン分子Sema4AのTh1/Th2反応制御機構の解明 2006 – 2007
- Th1/Th2バランス関連サイトカン遺伝子多型とうつ病発症・自殺企図との関連性 2005
- 樹状細胞とNKT細胞によるTh1/Th2サイトカインバランスの調節機構とその応用 2005 – 2006
- 新規ヒト腎炎モデルマウスの病態解析及びTh1/Th2転写制御による治療の試み 2005 – 2006
- Th1/Th2細胞分化・機能維持とクロマチンリモデリングに関する研究 2005 – 2007
- 好塩基球を介したTh1/Th2分化制御および感染に対する免疫監視機構の研究 2005 – 2006
- DNAM-1による樹状細胞の活性化とTh1/Th2バランスの制御機構 2005 – 2006
- Th1/Th2バランス制御を介した抗腫瘍免疫の誘導とそのメカニズムの解析 2005 – 2006
- Th1,Th2サイトカインの皮膚バリア機能に及ぼす影響 2004 – 2006
- 耳鼻咽喉科領域疾患におけるTh1,Th2,Tc1,Tc2細胞の検討 2004 – 2005
- 黄砂のTh1・Th2免疫系および経口免疫寛容に与える影響 2004 – 2006
- 抗原提示細胞を介したTh1/Th2細胞への分化・誘導制御機構と免疫細胞療法 2004 – 2007
- Th1/Th2分化におけるIL-12レセプターβ1遺伝子プロモーター機能の役割 2004 – 2005
- CD8α陽性樹状細胞の活性化とTh1/Th2バランスの制御機構 2004
- Th1/Th2細胞における皮膚ホーミングレセプター発現機序に関する研究 2003 – 2004
- クロマチンリモデリング選択的GATA-3変異体を用いたTh1/Th2バランス制御 2003
- 内分泌攪乱物質の免疫攪乱誘導による免疫毒性作用 2003 – 2004
- Th1/Th2病としての自己免疫疾患の病態解明と予防・治療 2003 – 2005
- サイトカイン抑制制御分子SOCSを利用したTh1・Th2バランスの改変 2003
- 転写因子AML1によるT細胞のTh1/Th2系列への振り分け制御機構の解明 2003
- 表皮におけるセラミド合成に対するTh1、Th2サイトカインの影響 2002 – 2003
- アトピー性皮膚炎の表皮細胞におけるTh1とTh2ケモカイン産生の制御について 2002 – 2003
- Decoy DNAによるTh1/Th2サイトカインの発現調節と免疫応答制御の検討 2002 – 2003
- Th1,Th2への分化に伴うE-セレクチン・リガンドと糖転移酵素の発現 2002 – 2003
- TH1・TH2バランスと自然免疫のクロストークにおける細胞内レドックスの役割 2002 – 2003
- 免疫学的生殖不全におけるNK・NKT細胞,Th1/Th2サイトカインの役割の解明 2002 – 2004
- Th1/Th2分化制御による慢性関節リウマチの分子標的療法開発への基礎的研究 2002 – 2003
- Th1/Th2細胞分化とクロマチンリモデリングに関する研究 2002 – 2004
- 内分泌攪乱物質のTh1/Th2免疫応答への影響と易感染性との関連 2002 – 2003
- サイトカインシグナル抑制分子SOCSを利用したTh1・Th2バランスの改変 2002 – 2003
- 樹状細胞によるTh1/Th2バランスの決定機構 2002
- 食餌制限によるTh1/Th2バランスの制御とその癌ワクチン療法への応用 2002 – 2004
- 炎症性骨吸収におけるヘルパーT細胞(Th1・Th2細胞)の役割 2002 – 2004
- 炎症性骨吸収におけるヘルパーT細胞(Th1・Th2細胞)の役割 2002 – 2004
- 妊娠時におけるTh1/Th2バランスに関する検討 2001
- バセドウ病モデルマウスの病態におけるTh1/Th2細胞・IL-5が果たす役割 2001
- 敗血症におけるTh1/Th2サイトカインの作用と調節機構の解明 2001 – 2002
- Th1/Th2細胞分化機構の制御によるがん免疫の賦活化 2001
- 口腔扁平上皮癌患者の放射線治療によるTh1/Th2バランスの変化について 2000 – 2001
- 侵襲時における末梢血Th1/Th2バランスの変動とサイトカイン動態に関する検討 2000 – 2001
- ヘルパーT細胞Th1/Th2バランスを制御する転写因子の検索 2000 – 2001
- マイクロアレイ法を用いたTh1,Th2型免疫応答の分類化と免疫疾患の病態解析 2000
- 卵巣癌患者末梢血幹細胞移植におけるTh1/Th2解析に関する研究 1999 – 2000
- 癌患者におけるHelper T CellのTh1/Th2バランスに関する研究 1999 – 2000
- 自己免疫疾患発症の分子機構の解明およびその制御:細胞表面分子群のTh1/Th2分化・誘導機構の解明とその制御 1999 – 2000
- 全身性エリテマトーデスにおけるTh1/Th2バランスの解析 1999 – 2002
- Th1/Th2細胞の活性化による微生物に対する感染防御能の増強効果 1999 – 2001
- Th1/Th2バランス制御法の癌免疫療法への応用 1999
- 間質性肺炎に於けるTh1/Th2細胞及びIL-2の役割の研究 1999 – 2000
- Th1/Th2バランス制御因子の遺伝子解析とその免疫疾患との関連性 1998 – 1999
- Th1/Th2細胞誘導を指標としたBRM感受性試験の開発と臨床応用 1998 – 2000
- Th1/Th2バランスからみたHAM発症分子機構の解明と治療法開発の基礎的検討 1998 – 1999
- シェーグレン症候群ににおけるTh1/Th2バランス異常の解析とその制御戦略 1998 – 1999
- 外科侵襲に対する免疫応答におけるTh1/Th2システムの証明 1997
- アトピー性皮膚炎患者のTh1-Th2バランス制御機構の解析 1997 – 1998
- 自己腫瘍特異的CTLのTCRVβレパトアとTh1/Th2サイトカイン産生能の解析 1997 – 2000
- IgA腎症病態発現へのTh1/Th2バランス制御の効果-若年好発症IgA腎症 (HIGA) マウスへのIL-12投与の検討- 1997 – 1998
- C型肝炎ウィルス特異的ヘルパーTクローン樹立と抗原依存性TH1・TH2分化の検討 1996 – 1997
- アトピー性皮膚炎患者におけるTh1-Th2バランス制御機構の解析 1996
- 遅延型過敏症を担うTh1,Th2細胞活性化における抗原提示細胞表面分子の役割 1996
- マウスでの抗原反復投与によるTH1/TH2不均衡の成立機序:マウスのアトピー性皮膚炎モデル作成とその検討 1996
- フローサイトメトリーによるTh1/Th2リンパ球の解析-猫条虫感染マウス系におけるエフェクター細胞の検討- 1996
- 炎症性皮膚疾患におけるTh1 Th2細胞へのdeviationを規定する因子の解析 1996
- AIDS粘膜ワクチン:Th1/Th2型細胞によるHIV特異的粘膜免疫の誘導 1996
- 自己免疫性心筋炎の発症における細胞間分子およびTh1/Th2サイトカンの役割の解明と抗接着分子療法による治療に関する臨床的・実験的検討 1995
- 寄生虫感染におけるTh1,Th2サブセット活性化機構の解明 1995 – 1996
- AIDS粘膜ワクチン:Th1/Th2型細胞によるHIV特異的粘膜免疫の誘導 1995
- 皮膚樹状細胞とアレルギー,Th_1/Th_2サイトカイン群と抗原提供能 1994
- 広東住血線虫感染マウスのTh1/Th2サイトカイン応答 1994 – 1995
- 腎細胞癌内TH1/TH2免疫調節機構、T細胞受容体解析および腎細胞癌免疫療法 1993 – 1994
-
Th17の発見とTh1/Th2パラダイムの修正
The classical T helper (Th)1 and Th2 CD4+ T cell effector paradigm has recently been challenged. Studies from various laboratories have shown the existence of a T cell subpopulation, dubbed Th17, not only distinct from Th1 and Th2, but a different pro-inflammatory Th-cell lineage.( Autoimmunity Reviews Review Autoimmune inflammation from the Th17 perspective Autoimmunity Reviews Volume 6, Issue 3, January 2007, Pages 169-175 無料要旨)
Helper T cells are CD4+ T lymphocytes that have an important role in determining the nature of the adaptive immune response. Since the 1980s, helper T cells have been classified in two major types, namely the Th1 or Th2 phenotype. Th1 cells promote cellular immunity, which is associated with anti-viral responses and tumour surveillance, whereas Th2 cells promote humoral responses to extracellular parasites and are involved in allergies. ‥ The two-dimensional Th1/Th2 paradigm has been a very successful foundation of immunology for the past 20 years. However, the recent discovery of Th17 cells confirmed earlier evidence that helper T cells may adopt phenotypes other than Th1 and Th2. (From the two-dimensional Th1 and Th2 phenotypes to high-dimensional models for gene regulation International Immunology, Volume 20, Issue 10, October 2008).
For almost two decades, the Th1/Th2 paradigm has offered a productive conceptual framework for investigating the pathogenesis of periodontitis. However, as with many other inflammatory diseases, the observed role of T-cell-mediated immunity in periodontitis did not readily fit this model. A new subset of CD4+ T-cells was recently discovered that explains many of the discrepancies in the classic Th1/Th2 model, and has been termed “Th17” based on its secretion of the novel pro-inflammatory cytokine IL-17. (A New Inflammatory Cytokine on the Block: Re-thinking Periodontal Disease and the Th1/Th2 Paradigm in the Context of Th17 Cells and IL-17 September 1, 2008 )
Th17細胞は粘膜免疫の維持に不可欠である一方で、それらの調節障害は自己免疫性炎症の病理発生に関与しています。標準的なTh1/Th2モデルとは異なるCD4+ヘルパーT細胞の3番目のサブセットの存在が最初に示唆されたのは、この炎症を引き起こす際のそれらの役割でした。元のモデルでは、Th1細胞は自己免疫の主なメディエーターであると考えられていました。しかし、それらの主要なTh1エフェクターサイトカインであるIFNγまたはそれらの活性化サイトカインであるIL-12が欠如すると、実験的自己免疫性脳脊髄炎(EAE)などの自己免疫性炎症のモデルが悪化することが明らかにされました。IL-12/IFNγではなくIL-23/IL-17系が、この炎症を媒介する主な経路として特定され、その後の研究では、この系に関連する新規のCD4+ヘルパーTサブセットとしてTh17細胞の特徴が明らかにされました。それ以来、Th17細胞は、関節リウマチ、乾癬、多発性硬化症、および炎症性腸疾患などの他の自己免疫疾患の進行に役割を果たすことが明らかにされています(ヘルパーT17細胞(Th17細胞)とは? ThermoFisher Scientific)
今となってはシンプルにすらみえるTh1/Th2分化の概念図に、Th17細胞という新しいサブセットが登場し、Th17細胞分化が制御性T細胞分化と相互背反的に成立することが示されるに至って、今や各T細胞サブセットの関連を、はるかに複雑な構図のなかで据え直す必要に迫られている(サイトカインネットワークのパラダイムシフト)
これまでぜんそくをはじめとした種々のアレルギー疾患は、Th1細胞とTh2細胞のバランスが崩れることが病態形成の引き金となる(Th1/Th2アンバランスモデル)と考えられてきましたが、本研究グループは、記憶Th細胞中の病原性を持った集団によりアレルギー疾患の病態が慢性化する(病原性記憶Th細胞亜集団疾患モデル)という新たなコンセプトを提唱しています(ぜんそくなどのアレルギー性気道炎症の慢性化機構を解明~難治性アレルギー疾患の新規治療薬開発に期待~ポイント 平成27年2月18日科学技術振興機構(JST)千葉大学)
- Th17 and Treg Cells Innovate the Th1/Th2 Concept and Allergy Research Blaser K (ed): T Cell Regulation in Allergy, Asthma and Atopic Skin Diseases. Chem Immunol Allergy. Basel, Karger, 2008, vol 94, pp 1-7(無料要旨)
- Kelso A, Troutt AB, Maraskovsky E, et al. Heterogeneity in lymphokine profiles of CD4+ and CD8+ T cells and clones activated in vivo and in vitro, Immunol. Rev., 1991, vol. 123 pg. 85
その他の参考
- Salivary gland tissue expression of interleukin-23 and interleukin-17 in Sjögren’s syndrome: Findings in humans and mice Cuong Q. Nguyen, Min H. Hu, Yi Li, Carol Stewart, Ammon B. Peck First published: 29 February 2008