オーガナイザーという概念についてのそもそも論
Spemannは、原口背唇部をオーガナイザー(発生過程全体を組織化する、発生の主催者といった意味合いがある)と呼んだのですが、原口背唇部をオーガナイザーと呼ぶことには、当時から強い異論がありました。Spemann自身1938年の著作で「私は現象の目覚ましさを表現しただけで、オーガナイザー説を唱えているわけではない」[1]と述べ、Spemann門下の2巨匠であるJohannes HoltfreterとVictor Hamburgerが1955年までの両生類の胚操作の研究を網羅的にまとめた総説[2]では、彼らは「仮にオーガナイザーと呼ぶことにする」と注意深く記載しており、Holtfreterは「オーガナイザーは名前のつけ損ない(misnomer)」[3]と言い切り、Hamburgerも1988年の名著「実験発生学が遺したもの:ハンス・シュペーマンとオーガナイザー」[4] の中で、オーガナイザーという考え方は再検討すべきだということを慎重な表現の中で幾度も述べています。
それにもかかわらず、オーガナイザーの名前と仮想の機能は神格化され、どの教科書にも事実として記載され、世界中に広まりました。Spemannが「オーガナイザーの発見」によって、1935年にノーベル賞を受賞したことの副作用もあったのかもしれません。
発生生物学の静かな革命 VOL.1 実験発生学とオーガナイザー 近藤寿人(JT生命誌研究館 顧問・表現ディレクター) JT生命誌研究館https://www.brh.co.jp/research/hisato_kondo/1/
オーガナイザーの定義
- 胚細胞の大掛かりな移動(原腸陥入)が起きている領域がある
- 別の胚(宿主胚)の他の場所に移植すると(例え不完全であったとしても)主に宿主胚の細胞から構成される2つ目の胚構造を生み出すという働きをする
- 2つ目の胚構造を構成する胚の部分としては、(1)頭部、(2)骨格や筋肉そのほかのもととしての中胚葉組織、(3)胴体の中心線に沿って伸びる、もう一つの中胚葉組織である脊索、(4)消化管のもとになる内胚葉組織など
発生生物学の静かな革命 VOL.1 実験発生学とオーガナイザー 近藤寿人(JT生命誌研究館 顧問・表現ディレクター) JT生命誌研究館https://www.brh.co.jp/research/hisato_kondo/1/
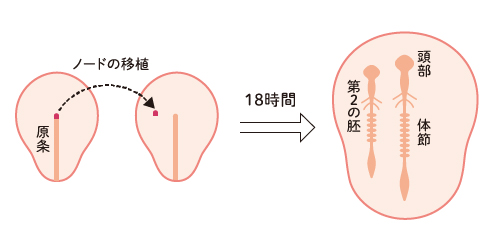
ニワトリのオーガナイザーは原始結節か
- 『他の脊椎動物の胚の発生も「オーガナイザー」という中核的な組織の働きに依存して発生するはずである』
- 哺乳類と鳥類の胚ではノード(ヘンゼン結節とも呼ばれる)といわれる小領域、魚類胚では胚盾と呼ばれる不均一な組織集団が、実験に基づく根拠があやふやな状況で「オーガナイザー」であると決め付けられてしまった
- 次のような図がまことしやかに教科書に描かれるように
このような鳥類胚を用いたノード移植実験は、私たちが実行するまで行われたことはなかったし(実際には、胚の培養技術を改善して初めて可能になった難しい実験)、この図のような結果は決して得られることはないのです。
発生生物学の静かな革命 VOL.1 実験発生学とオーガナイザー 近藤寿人(JT生命誌研究館 顧問・表現ディレクター) JT生命誌研究館 https://www.brh.co.jp/research/hisato_kondo/1/
- On the nature and function of organizers Alfonso Martinez Arias ORCID logo , Ben Steventon ORCID logo Author and article information Development (2018) 145 (5): dev159525. 09 March 2018 https://doi.org/10.1242/dev.159525
上のような解説がありましたが、ニワトリのオーガナイザーに関する総説論文
The avian organizer THOMAS BOETTGER1, HENDRIK KNOETGEN, LARS WITTLER and MICHAEL KESSEL* Int. J. Dev. Biol. 45: 281-287 (2001) https://ijdb.ehu.eus/article/pdf/11291858
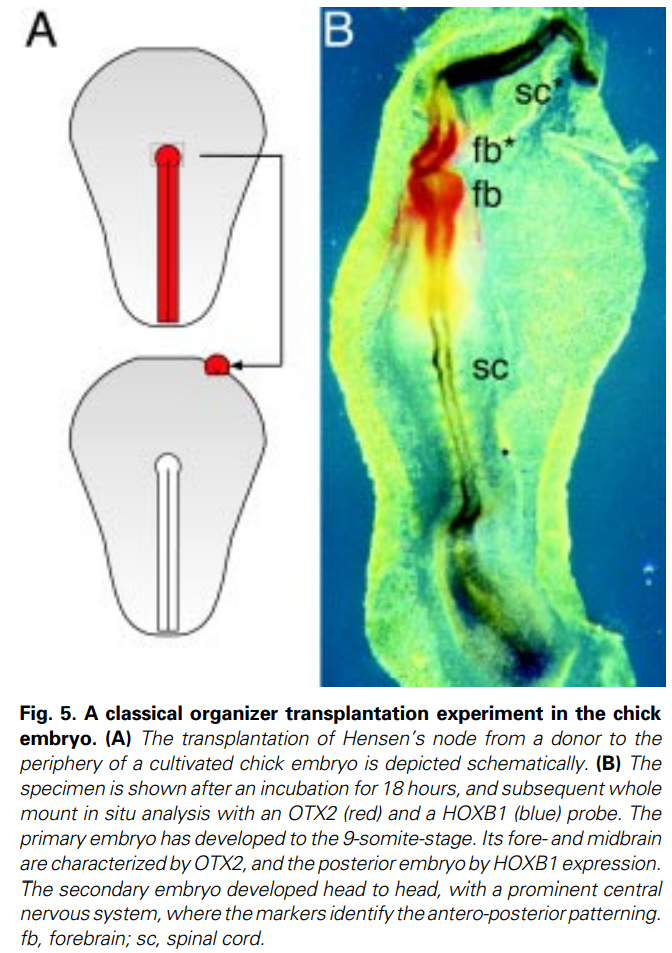
を見ると、「古典的な実験」として、ニワトリ胚の2次軸形成が紹介されていました。そのデータの原著、出典は明記されていません。著者が自分の研究室で行った未発表データなのでしょうか、それとも何かほかの論文からでしょうか。他の論文でも、2次軸が部分的に形成された例ならありましたが、移植された結節とは別に、一次軸から2次軸が枝分かれしているようです(下のFig.8)なぜ移植片が周囲の組織を誘導するのでなく、遠く離れた一次軸から2次軸を枝分かれさせるのでしょうか。本文にどう書いているかというと
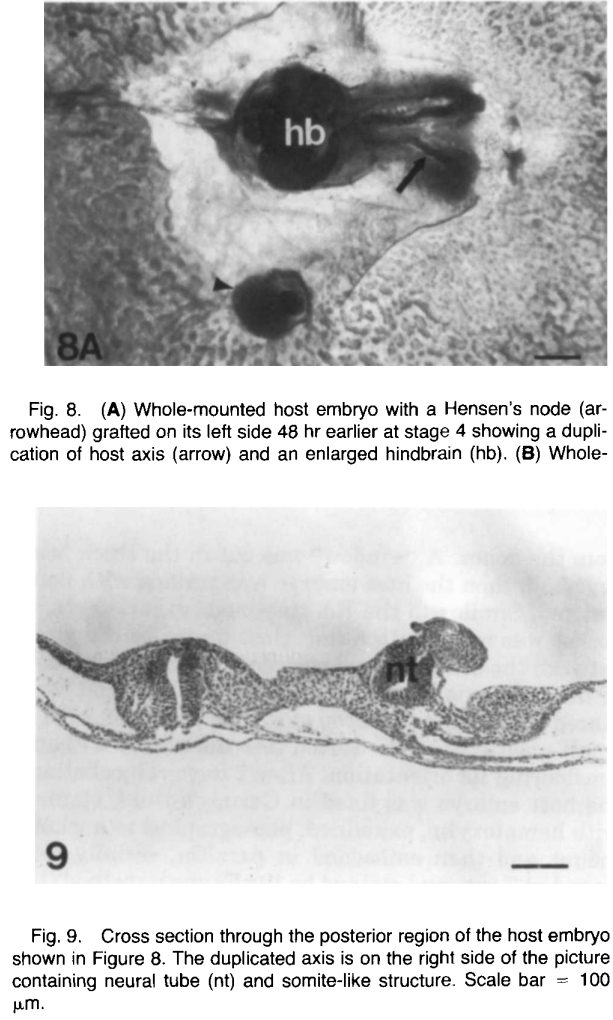
Inductive Effects of Grafted Hensen’s Node
Hensen’s node taken from stage 4 to stage 9 quail embryos (the majority was from stage 4) was transplanted to different sides of the axis of stage 4 to stage 6 chick embryos in the area pellucida. The graft differentiated into notochord, neural tissues, somites, and mesenchymal cells, and could also induce the associated host ectoderm to form neural tissue, as described previously (Gallera, 1971; Hornbruch et al., 1979). However, after examining the host embryos in detail, we found that a third axiswas formed in many host embryos besides the secondary axis formed by the grafted Hensen’s node itself (Fig. 8). The third axis was duplicated entirely from host tissue and usually contained a neural tube and somites (Fig. 9). As shown in Table 4, the stage 4 embryo was again the most sensitive to the inductive action of Hensen’s node. Like RA, Hensen’s node grafted on the left side or anterior side of the host axis could cause the duplication of the host axis most frequently (see Table 4). However, Hensen’s node produces more complicated inductive effects than RA. When Hensen’s node was grafted on the right side of the stage 4 host axis, it could also induce a third axis. Stage 5 embryos still responded to the inductive action of grafted Hensen’s node and yielded a third axis. Interestingly, if stage 5 embryos were used as the hosts, only those hosts that received a grafted Hensen’s node on the right side of their axis showed the duplication of the host axis. These results suggested that Hensen’s node might contain another inducing factor that acts like RA but in the opposite direction or might be more potent than the RA beads. However, if stage 6 embryos were used as hosts, grafted Hensen’s node failed to cause axial duplication in the hosts. When primitive streak taken from stage 4 to stage 6 quail embryos was transplanted to stage 4 chick blastoderms as a control experiment, none of the total 12 cases showed host axis duplication (Fig. 8).
DEVELOPMENTAL DYNAMICS 19514%151 (1992) Comparison of Hensen’s Node and Retinoic Acid in Secondary Axis Induction in the Early Chick Embryo YIPING CHEN AND MICHAEL SOLURSH
- Gallera, J. (1971) Primary induction in birds. Adv. Morphog. 9:149-180 https://www.sciencedirect.com/science/article/abs/pii/B978012028609650008X
- Hornbruch, A,, Summerbell, D., and Wolpert, L. (1979) Somite formation in the early chick embryo following grafts of Hensen’s node. J. Embryol. Exp. Morphol. 51:51-62. https://pubmed.ncbi.nlm.nih.gov/479748/
他にも本文や図はみられませんがグーグル検索で引っかかった図として、下のものがありました。
https://ars.els-cdn.com/content/image/1-s2.0-S0070215324000255-gr2.jpg

かなりややこしい状況です。確かに、上の日本語の総説のような完全な2次軸が形成されるというデータは見当たらないかもしれません。
Consensus.aiで「ニワトリ胚でHensen’s nodeを異所性に移植すると、完全な体軸が誘導され、ホスト由来の神経組織を含む完全な胚軸が形成されるか?」と訊いてみると、100%YESの回答がきました。しかし根拠として挙げられてている論文を見ると、どうも全然根拠になっていないようです。生成AIは、適当に答えをでっちあげている可能性があります。生成AIはネット上の文書をインプットしているので、ネット上の文書が間違っていると、それらを統合した出力も当然間違っていることになります。
完全な体軸の誘導:Hensen’s nodeを異所性に移植すると、ホスト由来の神経組織を含む完全な体軸が形成されることが示されています12456789。
神経誘導の能力:Hensen’s nodeは、異所性に移植された場合に新しい神経軸を誘導する能力があり、特に若い段階のnodeが強い誘導能力を持つことが確認されています2357。
ホストとドナーの寄与:異所性に移植されたHensen’s nodeは、ホスト由来の神経組織を誘導し、ドナー由来の組織と融合することなく独立して発達することが観察されています469。
年齢の影響:Hensen’s nodeの年齢が神経誘導の結果に影響を与え、若いnodeは前部および後部の神経系を誘導し、年齢が進むと後部の神経系のみを誘導する傾向があります27。
分子メカニズム:Hensen’s nodeによる神経誘導には、特定の転写因子や分泌分子が関与しており、これらが胚全体に拡散して神経板の形成を誘導することが示唆されています358。
結論 Hensen’s nodeを異所性に移植すると、ホスト由来の神経組織を含む完全な体軸が形成されることが複数の研究で確認されています。この能力は、特に若い段階のnodeで顕著であり、神経誘導には特定の分子メ THOMAS BOETTGER1, HENDRIK KNOETGEN, LARS WITTLER THOMAS BOETTGER1, HENDRIK KNOETGEN, LARS WITTLER R
- The avian organizer THOMAS BOETTGER1, HENDRIK KNOETGEN, LARS WITTLER and MICHAEL KESSEL* Int. J. Dev. Biol. 45: 281-287 (2001) https://ijdb.ehu.eus/article/pdf/11291858
- Restoration of the organizer after radical ablation of Hensen’s node and the anterior primitive streak in the chick embryo Delphine Psychoyos, Claudio D. Stern Author and article information Development (1996) 122 (10): 3263–3273. 01 October 1996 https://doi.org/10.1242/dev.122.10.3263 https://journals.biologists.com/dev/article/122/10/3263/38976/Restoration-of-the-organizer-after-radical
- Axis development in avian embryos: the ability of Hensen’s node to self-differentiate, as analyzed with heterochronic grafting experiments Takayuki Inagaki, Gary C. Schoenwolf Anat Embryol (1993) 188:1-11 Type-3 experiments. Ten cases of ectopic embryos were obtained from the 13 host blastoderms. In such ectopic embryos, the transplanted nodes self-differentiated into notochord, somites, mesenchyme, and endoderm; the host’s ectoderm typically responded to the grafted cells by forming a neural tube, which often had a morphology characteristic of that of rostral brain levels (i.e., forebrain and midbrain (Figs. 10, 11). https://link.springer.com/article/10.1007/BF00191446 この写真をみても外観を見る限り、きれいな2次軸が形成されるというほどではないようです。組織切片をみて、それぞれの組織への分化が確認できたとのことのようです。
- Formation of ectopic neurepithelium in chick blastoderms: Age-related capacities for induction and self-differentiation following transplantation of quail Hensen’s nodes Mark S. Dias, Dr. Gary C. Schoenwolf First published: December 1990 https://doi.org/10.1002/ar.1092280410 本文有料
- Neural induction and regionalisation in the chick embryo. Yes 若いヘンゼン結節 (ステージ 2 ~ 4) は前部神経系と後部神経系の両方を誘導しますが、古い結節 (ステージ 5 ~ 6) は後部神経系のみを生成します。 Development Kg Storey et al. 261 Citations 1992
- Multiple neural inductions in area opaca by grafts of the Hensen’s node do not retard host chick embryo development. Yes ヘンゼン結節の移植による不透明領域での複数の神経誘導は、宿主のニワトリ胚の発育を遅らせない。 Indian journal of experimental biology G. Guttikar et al. 3 Citations 1993
- Neural induction and regionalisation by different subpopulations of cells in Hensen’s node. Unknown 神経誘導能はヘンゼン結節の内側部の胚盤葉上層と中内胚葉に局在しているが、後外側部の深部にはこの能はない。 Development Kate G. Storey et al. 56 Citations 1995
灰色新月環 gray crescent
http://digfir-published.macmillanusa.com/pol2e/asset/img_ch38/c38_fig01.html
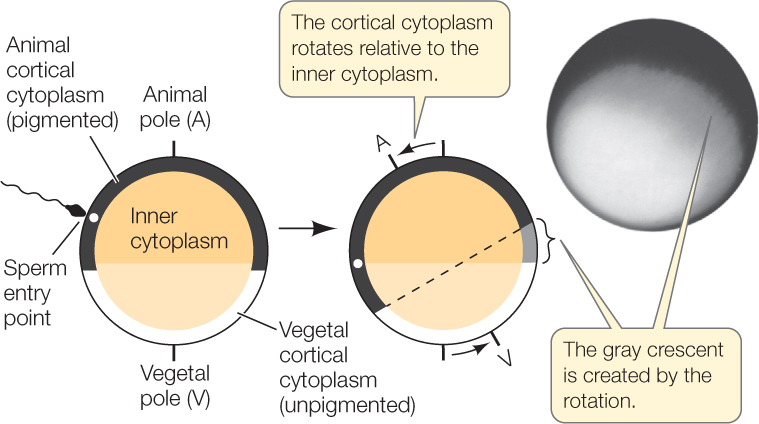
アフリカツメガエルの原腸陥入
灰色新月環は下の動画だと良くわかりません。
Xenopus laevis; gastrulation through neurulation Oxford Academic (Oxford University Press) チャンネル登録者数 15.6万人
シュペーマンのオーガナイザーの定義
The organizer is a region of the gastrula embryo that can induce the formation of a second neural axis when transplanted to an undifferentiated region of a host gastrula stage embryo (1). https://pmc.ncbi.nlm.nih.gov/articles/PMC27625
原図(A~C) Hamburger, V. 1988. The Heritage of Experimental Embryology: Hans Spemann and the Organizer. Oxford University Press, Oxford. 図引用元:Scotte Gilbert Developmental Biology. 6th edition. Axis Formation in Amphibians: The Phenomenon of the Organizer https://www.ncbi.nlm.nih.gov/books/NBK10101/
シュペーマンのオーガナイザーの今昔に関しては、International Journal of Developmental Biologyの特集号が非常に有益だと思います。2001年の特集号なので、今となってはそこからさらに24年の歳月が経過してはいますが、2001年当時の理解がわかります。Scott Gilbertの総説は非常に読み応えがありました。さすが大著のベストセラーの教科書を書いているだけのことはあります。読ませます。
- The International Journal of Developmental Biology Volume 45 > Issue 1 (Special Issue) Cover Vol. 45 N. 1 The Spemann-Mangold Organizer Edited by: Eddy DeRobertis and Juan Aréchaga https://ijdb.ehu.eus/issue/45/1
シュペーマンのオーガナイザーとして同定されたシグナル分子は驚くほど多数あります。オーガナイザーとして積極的に働く分子があるというよりも、デフォルトでは尾部を形成するシグナルとしてBMPシグナルとWntシグナルがあり、それらに対する阻害因子が頭部を形成するという図式になっています。そのため、オーガナイザー分子は、BMPシグナリングの阻害因子とWntシグナリングの阻害因子とに大別されます。BMPはTGF-βスーパーファミリーに属しています。
シグナル分子がやまのように出てきてかなり混乱させられるのでまずは主要なシグナル伝達経路を復習しておきます。
TGF-βファミリー分子のシグナル経路:R-SmadとCo-Smad
ちなみにTGF-βスーパーファミリーは、3つのサブファミリーに分かれ、TGF-βファミリー、アクチビンファミリー、BMP(bone morphogenetic protein)ファミリーがあります。TGF-βスーパーファミリーは、Smadがエフェクターとなる共通するシグナル経路によって特徴づけられています。まず受容体ですが、細胞膜上に存在する「2型受容体」と「1型受容体」の2種類の受容体があります。TGF-βが結合した2型受容体は1型受容体と複合体を形成します。2型受容体はセリン・スレオニンキナーゼで、1型受容体をリン酸化します。1型受容体もセリン・スレオニンキナーゼ型受容体でリン酸化によりキナーゼ活性をもち、R-Smadをリン酸化します。R-Smadにはいくつかの種類があります。Smad2、Smad3は、TGF-βやアクチビンのシグナルを伝えるエフェクターです。Smad1、Smad5、Smad8は、BMPのシグナルを伝えるエフェクターです。リン酸化されたR-Smadは、Co-Smadと呼ばれるSmad4と結合して複合体をつくります。R-SmadとCo-Smadの複合体は、細胞質から核に移行し、標的遺伝子の転写調節領域に存在するSBE(Smad binding element)に結合することにより、ほかの転写因子などと協同して標的遺伝子の発現を制御します。ほかに、R-SmadやCo-Smadの働きを抑制するI-Smadとしては、Smad6とSmad7があります。
- https://www.md.tsukuba.ac.jp/epatho/privacy.html
上の文章による説明にピッタリ合う模式図が、下の論文にありました。
https://www.researchgate.net/publication/7654580_Non-smad_TGF-beta_signals
Non-Smad TGF-β signals J Cell Sci (2005) 118 (16): 3573–3584. https://journals.biologists.com/jcs/article/118/16/3573/28415/Non-Smad-TGF-signals

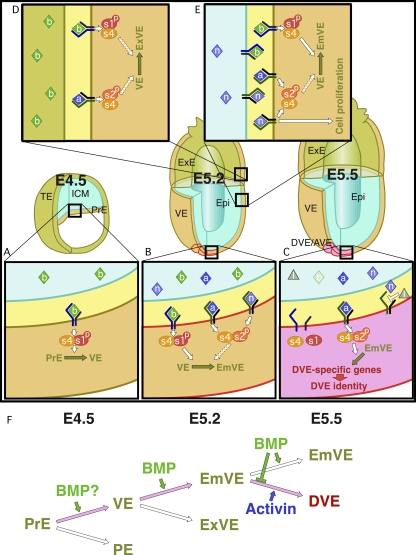
- Antagonism between Smad1 and Smad2 signaling determines the site of distal visceral endoderm formation in the mouse embryo J Cell Biol. 2009 Jan 26;184(2):323–334. doi: 10.1083/jcb.200808044 https://pmc.ncbi.nlm.nih.gov/articles/PMC2654303/
nodalシグナリング:カノニカル経路と非カノニカル経路
nodalはTGF-βファミリーに属するので、カノニカル経路としてはSmadの活性化があります。
Schematic Overview of Nodal and Cripto-1 Signaling Pathways. A) Canonical Nodal signaling using Cripto-1 as a co-receptor. Nodal can also activate this pathway, albeit less efficiently, independently of Cripto-1. B) Cripto-1 Nodal-independent signaling via binding with Glypican-1 and activating src/Ras/raf/PI3K downstream signaling. C) Possible signaling of Cripto-1 through binding to beta-integrins and signaling via FAK-dependent signaling pathway.
2011 Sep 19. Published in final edited form as: Breast Dis. 2008;29:91–103. doi: 10.3233/bd-2008-29110 Emerging Roles of Nodal and Cripto-1: From Embryogenesis to Breast Cancer Progression https://pmc.ncbi.nlm.nih.gov/articles/PMC3175751/#ref-list1
Wnt経路
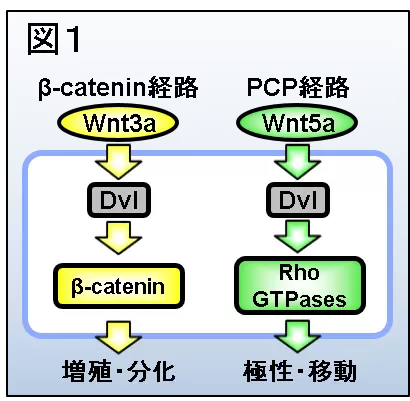
Wntの細胞内情報伝達機構にはいくつかの種類があります。カノニカル経路といわれるのはβ-カテニン経路です。
- 細胞はWnt-3aファミリーの分子を受容すると、その情報を細胞内情報伝達分子Dishevelled(Dvl)タンパク質へと伝えます(図1左)。これにより活性化したDvlはβ-catenin経路を活性化します。
- 細胞がWnt-5aファミリーの分子を受容した場合も、Dvlが活性化しますが、Wnt-5aの下流で活性化したDvlはPCP経路を活性化します。
- 細胞外情報伝達分子Wntは、動物の個体発生と恒常性の維持において、細胞の増殖・分化・極性・移動を制御します。
発癌に関与する細胞内情報伝達分子Dvlの新規調節機構を解明 https://www.tmd.ac.jp/mri/press/detail/id=58961
- Wntシグナル研究の歴史と展望~その足跡と未来 2020年8月25日 Journal of Japanese Biochemical Society 92(4): 498-516 (2020) doi:10.14952/SEIKAGAKU.2020.920498 https://seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2020.920498/data/index.html Int1のトランスジェニックマウスが生後6か月で乳がんを発症することも示され18),Int1は原がん遺伝子であると位置づけられるようになった.さらに,1987年にInt1はショウジョウバエのDNAにも存在することが判明し,その塩基配列が決定され,Drosophila int1(Dint-1)は,同年に塩基配列が報告されたショウジョウバエのwinglessと同一であることが決定した19, 20).ここで初めて,無脊椎動物の発生に関わる遺伝子と,哺乳動物のがんに関わる遺伝子が高い相同性(アミノ酸レベルで53%同一)を有することが明らかになった.‥ Int1欠損マウスは中脳と小脳の発生に異常が認められた.‥ MMTVプロウイルスを挿入することにより,活性化される遺伝子‥ Int1に類似している遺伝子群だけをWnt(Wingless-related integration site)と呼ぶようになった28).
Canonical and Non-Canonical Wnt Signaling Generates Molecular and Cellular Asymmetries to Establish Embryonic Axes J. Dev. Biol. 2024, 12(3), 20; https://doi.org/10.3390/jdb12030020 MDPI誌 https://www.mdpi.com/2221-3759/12/3/20
BMP(骨形成因子)の阻害因子
BMPはTGF-βスーパーファミリー(TGF-β、BMPs、Activin)のなかのサブファミリーの一つです。
Noggin
BMP4に高親和性で結合し、その活性を阻害する
- Expression cloning of noggin, a new dorsalizing factor localized to the Spemann organizer in Xenopus embryos W C Smith 1, R M Harland Cell 1992 Sep 4;70(5):829-40. doi: 10.1016/0092-8674(92)90316-5. 要旨より zygotic transcripts are initially restricted to the presumptive dorsal mesoderm and reach their highest levels at the gastrula stage in the dorsal lip of the blastopore (Spemann organizer). In the neurula, noggin is transcribed in the notochord and prechordal mesoderm. The activity of exogenous noggin RNA in embryonic axis induction
最初の論文は1992年のセルペーパーでした。
この論文を読むとUV照射により腹側化したカエル卵にnoggin mRNAを注入したときに体軸ができたというデータを見せていますが、単に腹側に打ったら2次軸ができたというデータはありません。UV照射が不完全であればもともとの1次軸ができてしまうので、この実験は結構微妙な実験だと思います。UV照射・mRNA無しの実験結果で5匹の胚を示していますが、容易にバイアスがかかりそう。
2次軸はできるのでしょうか、できないのでしょうか?Consensusに訊いてみました。
- ノギンmRNAの腹側注入による部分的な二次軸形成:ノギンmRNAを腹側に注入すると、部分的な二次軸が形成されるが、頭部構造は形成されない
- 。ノギンmRNAの他の部位への注入による完全な二次軸形成:ノギンmRNAを他の部位(例えば、動物極近く、辺縁帯、植物極)に注入すると、完全な二次軸が形成される。
- ノギンのBMPシグナル抑制機能:ノギンはBMPシグナルを抑制することで二次軸を誘導する。
- ノギンと他の因子の相互作用:ノギンは他の因子(例えば、Nodal関連遺伝子やWntシグナル)と協力して完全な二次軸を形成する。
consensus.app
- Ectopic expression of Xenopus noggin RNA induces complete secondary body axes in embryos of the direct developing frog Eleutherodactylus coqui Development Genes and Evolution Volume 210, pages 21–27, (2000) https://link.springer.com/article/10.1007/PL00008184
Chordin
- Xenopus chordin: A Novel Dorsalizing Factor Activated by Organizer-Specific Homeobox Genes Yoshiki Sasai 1, Bin Lu 1, Herbert Steinbeisser 1, Douglas Geissert 1, Linda K Gont 1, Eddy M De Robertis Cell. 1994 Dec 2;79(5):779–790. doi: 10.1016/0092-8674(94)90068-x https://pmc.ncbi.nlm.nih.gov/articles/PMC3082463/ (無料著者原稿)Seventy years after the organizer experiment (Spemann and Mangold, 1924), the quest to isolate the factors released by the organizing centers to recruit neighboring cell continues. 要旨 Microinjection of chordin mRNA induces twinned axes
最初の論文は1994年のセル論文でした。
Follistatin
- Follistatin, an antagonist of activin, is expressed in the Spemann organizer and displays direct neuralizing activity A Hemmati-Brivanlou 1, O G Kelly, D A Melton Cell . 1994 Apr 22;77(2):283-95. doi: 10.1016/0092-8674(94)90320-4.
最初の報告は1994年のCellペーパーでした。
- フォリスタチンmRNAの注入は、ゼノパス胚において異常な発達を引き起こし、特に脊索形成に影響を与えることが示されていますが、二次軸形成を直接誘導するという明確な証拠は提供されていません。他のmRNA(Xwnt-8、Wnt-1、Activin、xFKBP1Bなど)は、二次軸形成を誘導する能力を持つことが示されています。(Consensus.ai)
実はBMP阻害因子であるNoggin, Chordin, Follistatinは頭部を含む完全な2次軸は形成しなかったんですね。つまり他のシグナルと協同して働くということのようです。
Cerberus
- Protein Related to DAN and Cerberus Is a Bone Morphogenetic Protein Antagonist That Participates in Ovarian Paracrine Regulation* Journal of Biological Chemistry Volume 279, Issue 22p23134-23141May 2004 https://www.jbc.org/article/S0021-9258(20)66678-9/fulltext
Wntシグナル経路の阻害因子
Cerberus
CerberusはWntシグナルを阻害するだけでなく、BMPの阻害やnodalシグナルの阻害もします。
Figrure 4. Model of the formation and function of anterior endoderm in Xenopus head development.
The head inducer Cerberus is a multifunctional antagonist of Nodal, BMP and Wnt signals Nature volume 397, pages707–710 (1999)
https://www.nature.com/articles/17820
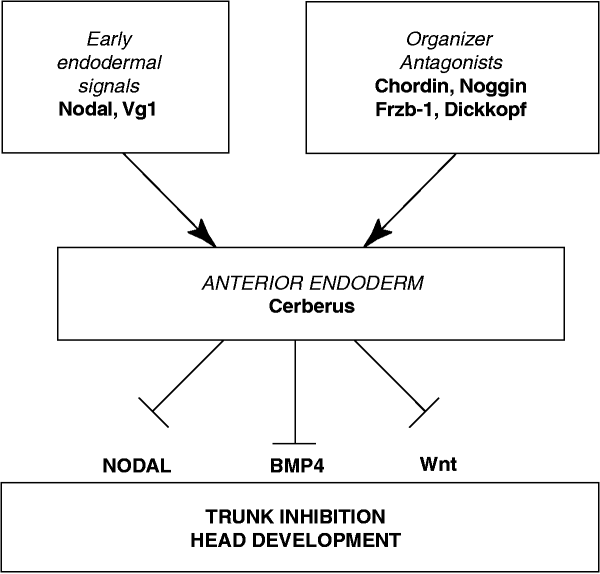
cerberusはWntリガンドと直接結合することによりWntシグナリングを抑制します。
https://www.sciencedirect.com/topics/biochemistry-genetics-and-molecular-biology/cerberus-protein

- Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemann’s organizer Tewis Bouwmeester, Sung-Hyun Kim, Yoshiki Sasai, Bin Lu & Eddy M. De Robertis Nature volume 382, pages595–601 (1996) 15 August 1996.
最初の報告は1996年のネイチャー論文でした。
- Head organizer: Cerberus and IGF cooperate in brain induction in Xenopus embryos Yagmur Azbazdar 1, Edgar M Pera 2, Edward M De Robertis 3 Cells Dev . 2023 Dec 16:203897. doi: 10.1016/j.cdev.2023.203897. イントロダクションより Cerberus induces small head structures containing brain, eyes, olfactory placodes, and sometimes a second heart, that are devoid of trunk-tail structures and posterior neural tissue.
- Molecular link in the sequential induction of the Spemann organizer: direct activation of the cerberus gene by Xlim-1, Xotx2, Mix.1, and Siamois, immediately downstream from Nodal and Wnt signaling Developmental Biology Volume 257, Issue 1, 1 May 2003, Pages 190-204
- Expression of the mouse cerberus-related gene, Cerr1, suggests a role in anterior neural induction and somitogenesis William Shawlot 1, Jian Min Deng 1, Richard R Behringer 1,* Proc Natl Acad Sci U S A. 1998 May 26;95(11):6198–6203. doi: 10.1073/pnas.95.11.6198
Dickkopf (Dkk)
- Head inducer Dickkopf-1 is a ligand for Wnt coreceptor LRP6 Current Biology Volume 11, Issue 12, 26 June 2001, Pages 951-961 The Wnt family of secreted growth factors initiates signaling via the Frizzled (Fz) receptor and its candidate coreceptor, LDL receptor-related protein 6 (LRP6), presumably through Fz-LRP6 complex formation induced by Wnt. ‥ Dkk-1 is a high-affinity ligand for LRP6 and inhibits Wnt signaling by preventing Fz-LRP6 complex formation induced by Wnt. Dkk-1 binds neither Wnt nor Fz, nor does it affect Wnt-Fz interaction. https://www.sciencedirect.com/science/article/pii/S0960982201002901
Frzb (Frizzled-related protein)
- Genes Dev. 1999 Sep 1;13(17):2328–2336. doi: 10.1101/gad.13.17.2328 Axis determination by inhibition of Wnt signaling in Xenopus Keiji Itoh 1, Sergei Y Sokol 1,1 we interfered with Wnt signaling in the embryo using the extracellular domain of Xenopus Frizzled 8 (ECD8), which blocks Wnt-dependent activation of a target gene in Xenopus ectodermal explants. Expression of ECD8 in ventral blastomeres resulted in formation of secondary axes この論文は内在的な分子の話ではなく、Wntシグナリングを阻害したときの効果(2次軸形成)をみるための実験
nodalの阻害因子
cerberus
cerberusは、Wntの阻害因子でもあり、BMP(TGF-βスーパーファミリー)の阻害因子でもありますが、nodal(TGF-βスーパーファミリー)の阻害因子でもあります。多才ですね。
Human Cerberus Prevents Nodal-Receptor Binding, Inhibits Nodal Signaling, and Suppresses Nodal-Mediated Phenotypes PLOS ONE January 201510(1):e0114954 DOI:10.1371/journal.pone.0114954 SourcePubMed LicenseCC BY 4.0 https://www.researchgate.net/publication/271219306_Human_Cerberus_Prevents_Nodal-Receptor_Binding_Inhibits_Nodal_Signaling_and_Suppresses_Nodal-Mediated_Phenotypes
![Proposed mechanism of Cerberus inhibition. (A) Structure of BMP9-ALK1-ACTRIIB complex [37]. The disulfide linked BMP9 homodimer (center, orange) simultaneously binds the extracellular domains of the type I Activin receptor-like kinase ALK1 (light blue) and the type II Activin receptor kinase ACTRIIB (dark blue). (B) Molecular model of Nodal-receptor interactions based on the BMP9-ALK1-ACTRIIB structure. The disulfide linked Nodal homodimer (center, orange) binds the extracellular domains of the type I Activin receptor-like kinase ALK4 (light blue) and the type II Activin receptor kinase ACTRIIB or BMPRII (dark blue), likely using canonical interaction surfaces (yellow lined circles). (C-D) Cerberus forms a stable homodimer in solution [58], binds Nodal and prevents binding of Nodal to both receptors. Thus Cerberus blocks simultaneously the ALK4 interaction surface (light blue circle) and the ACTRIIB interaction surface (dark blue circle). We propose that one Cerberus protomer could block interaction surfaces within one Nodal protomer (C) or two protomers (D). We expect the overall stoichiometry of this complex to be 2:2.](https://www.researchgate.net/profile/Washington-Mutatu/publication/271219306/figure/fig11/AS:341770715058184@1458495896759/Proposed-mechanism-of-Cerberus-inhibition-A-Structure-of-BMP9-ALK1-ACTRIIB-complex.png)
https://www.researchgate.net/figure/The-Nodal-signaling-pathway-Genetic-interaction-and-co-Immunoprecipitation-co-IP_fig10_271219306

- Nodal antagonists regulate formation of the anteroposterior axis of the mouse embryo. Yamamoto M, Saijoh Y, Perea-Gomez A, Shawlot W, Behringer RR, Ang SL, Hamada H, Meno C. Nature. 2004;428:387–392. doi: 10.1038/nature02418. Abstract Patterning of the mouse embryo along the anteroposterior axis during body plan development requires migration of the distal visceral endoderm (DVE) towards the future anterior side by a mechanism that has remained unknown. Here we show that Nodal signalling and the regionalization of its antagonists are required for normal migration of the DVE. Whereas Nodal signalling provides the driving force for DVE migration by stimulating the proliferation of visceral endoderm cells, the antagonists Lefty1 and Cerl determine the direction of migration by asymmetrically inhibiting Nodal activity on the future anterior side. https://www.nature.com/articles/nature02418 本文読めた
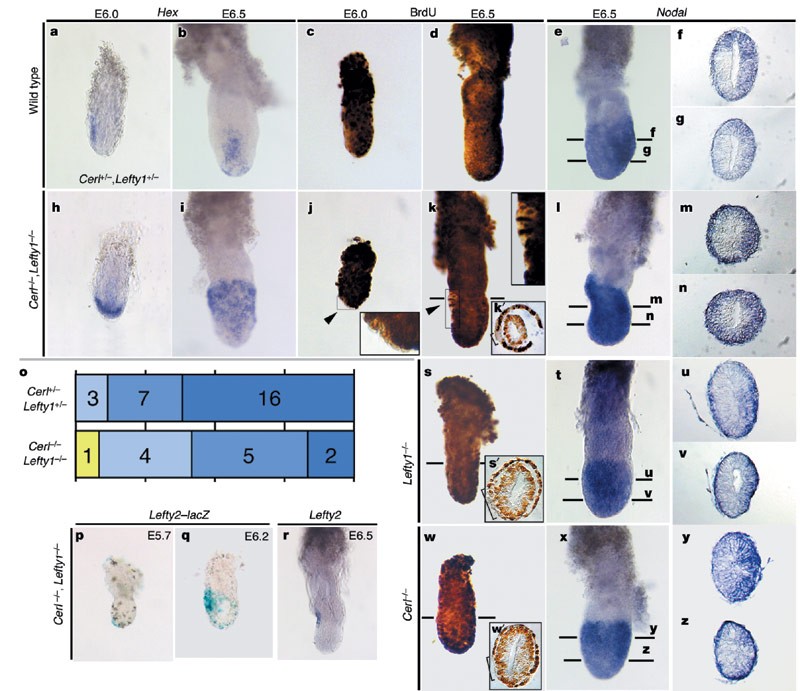 To confirm the role of Nodal antagonists in A–P patterning, we analysed mutant mice lacking Lefty1 or Cerl. Lefty1-/- (ref. 13) and Cerl-/- (refs 6, 14, 15) embryos undergo normal gastrulation, whereas Cerl-/-,Lefty1-/- embryos develop multiple primitive streaks8, suggestive of functional redundancy between Lefty1 and Cerl. We first examined expression of the AVE marker Hex (Fig. 5a, b, h, i). The Hex expression domain was expanded slightly in Cerl-/- and Lefty1-/- mutants (data not shown) and markedly in the Cerl-/-,Lefty1-/- double mutant (Fig. 5b, i), in which cells positive for Hex transcripts occupied the entire anterior half of the VE at E6.5 (n = 4). These results reveal that the Nodal antagonists restrict the size or location of the AVE, perhaps by inhibiting the proliferation of AVE cells.
To confirm the role of Nodal antagonists in A–P patterning, we analysed mutant mice lacking Lefty1 or Cerl. Lefty1-/- (ref. 13) and Cerl-/- (refs 6, 14, 15) embryos undergo normal gastrulation, whereas Cerl-/-,Lefty1-/- embryos develop multiple primitive streaks8, suggestive of functional redundancy between Lefty1 and Cerl. We first examined expression of the AVE marker Hex (Fig. 5a, b, h, i). The Hex expression domain was expanded slightly in Cerl-/- and Lefty1-/- mutants (data not shown) and markedly in the Cerl-/-,Lefty1-/- double mutant (Fig. 5b, i), in which cells positive for Hex transcripts occupied the entire anterior half of the VE at E6.5 (n = 4). These results reveal that the Nodal antagonists restrict the size or location of the AVE, perhaps by inhibiting the proliferation of AVE cells.
nodalはTGF-βファミリーの一員です。cereberusとLeftyはnodalの阻害因子として働きます。ちなみにcereberusはWntも阻害します。
a Following implantation, Nodal is expressed throughout the epiblast and plays a role in specifying the underlying distal visceral endoderm (DVE). The DVE in response secretes Nodal inhibitors Cerberus and Lefty, generating a posterior-distal gradient of Nodal and forming one of the earliest patterns in the embryo. ExE extra-embryonic ectoderm, VE visceral endoderm, P-D proximal-distal. b The DVE migrates anteriorly to become the anterior visceral endoderm (AVE) concentrating Nodal further to the proximal and newly formed posterior axis by continued secretion of Nodal inhibitors. A-P anterior-posterior. c In a process that requires high levels of Nodal, gastrulation begins with formation of the early primitive streak. Epiblast cells undergo EMT and migrate with subsequent differentiation into mesoderm and endoderm lineages. Inset: Pro-Nodal expressed in the epiblast upregulates Furin, Pace4, and BMP4 within the ExE. Furin and Pace4 subsequently process pro-Nodal and BMP4 stimulates Wnt3 signaling in the epiblast. Wnt3 acts on a Nodal proximal epiblast enhancer to increase Nodal expression. Nodal also amplifies its own signal through an enhancer region in intron 1 stimulated by Smad2/3-FoxH1, collectively increasing levels of Nodal in the proximal epiblast.
Plasticity underlies tumor progression: Role of Nodal signaling March 2016 Cancer and Metastasis Reviews 35(1) DOI:10.1007/s10555-016-9605-5 https://www.researchgate.net/figure/Role-of-Nodal-in-the-early-mouse-embryo-a-Following-implantation-Nodal-is-expressed_fig8_297661953

Epiblastの細胞がnodalを発現して分泌し、distal visceral endoderm (DVE)に働きかけます。するとDVEではnodalの阻害因子であるcerberus とLeftyを発現して、近くの細胞でのnodalの発現を抑制します。DVEは移動してAVEになりますが、するとAVE側のepiblastにおいてもnodalの発現は抑制されるので、AVEの反対側でnodalの発現が高いという状況が生まれます。このnodalシグナルがprimitive streakを引き起こすと考えられています。
- Nodal signaling from the visceral endoderm is required to maintain Nodal gene expression in the epiblast and drive AVE migration Amit Kumar 1,†, Margaret Lualdi 2, George T Lyozin 3, Prashant Sharma 1, Jadranka Loncarek 1, Xin-Yuan Fu 4, Michael R Kuehn Dev Biol. Author manuscript; available in PMC: 2016 Apr 1. Published in final edited form as: Dev Biol. 2014 Dec 20;400(1):1–9. doi: 10.1016/j.ydbio.2014.12.016 https://pmc.ncbi.nlm.nih.gov/articles/PMC4806383/ 要旨より The Nodal gene is expressed in both the VE and in the pluripotent epiblast, which gives rise to the germ layers. Previous findings have provided conflicting evidence as to the relative importance of Nodal signaling from the epiblast vs. VE for AP patterning.
その他
Goosecoid
アクチビン、Vg1、Xwnt-8、ノギンなどは、それぞれの細胞表面受容体を介して異なる細胞内シグナル伝達経路を開始し、最終的にgoosecoid (gsc) 遺伝子の発現を誘導
- Molecular Nature of Spemann’s Organizer: the Role of the Xenopus Homeobox Gene goosecoid Ken W Y Cho 1,*, Bruce Blumberg 1, Herbert Steinbeisser 1, Eddy M De Robertis 1 Cell. 1991 Dec 20;67(6):1111–1120. doi: 10.1016/0092-8674(91)90288-a https://pmc.ncbi.nlm.nih.gov/articles/PMC3102583/ 著者原稿無料PDF https://linkinghub.elsevier.com/retrieve/pii/009286749190288A 本文有料
goosecoid (gsc)がオーガナイザーの正体なのだとしたら、gsc遺伝子をノックアウトすれば、体軸を形成できないはずと予想されますが、予想に反してgscノックアウトマウスの表現型は、それほど大きくなかったようです。不思議です。
- Nasal and pharyngeal abnormalities caused by the mouse goosecoid gene mutation Biochem Biophys Res Commun . 1997 Apr 7;233(1):161-5. doi: 10.1006/bbrc.1997.6315. https://pubmed.ncbi.nlm.nih.gov/9144415/
Lim-1
- Role of the LIM class homeodomain protein Xlim-1 in neural and muscle induction by the Spemann organizer in Xenopus M Taira 1, H Otani, J P Saint-Jeannet, I B Dawid Nature . 1994 Dec 15;372(6507):677-9. doi: 10.1038/372677a0. https://www.nature.com/articles/372677a0
- Requirement for Lim1 in head-organizer function W Shawlot 1, R R Behringer Nature 1995 Mar 30;374(6521):425-30. doi: 10.1038/374425a0. Embryos homozygous for the null allele lacked anterior head structures but the remaining body axis developed normally.
カエルで発見されマウスでも役割が確認され、それぞれ1994年、1995年のネイチャー誌に報告されました。
- Functional domains of the LIM homeodomain protein Xlim-1 involved in negative regulation, transactivation, and axis formation in Xenopus embryos I Hiratani 1, T Mochizuki, N Tochimoto, M Taira Dev Biol . 2001 Jan 15;229(2):456-67. doi: 10.1006/dbio.2000.9986.
シュペーマンのオーガナイザーを誘導するシグナル
アクチビン、Vg1、Xwnt-8、ノギンがそれぞれの細胞表面受容体を介して異なる細胞内シグナル伝達経路を開始し、最終的にgoosecoid (gsc) 遺伝子の発現を誘導
Wntシグナル経路
Siamois Siamois遺伝子はWntシグナル経路のターゲットで、オーガナイザー活性を誘導する。Siamoisは中胚葉誘導を経ずにオーガナイザー特異的遺伝子を活性化し、背側化シグナルを生み出す
Xenopus nodal-related (Xnr) : Xnr因子はアクチビン様分子によって誘導され頭部オーガナイザーを形成。
TGF-β経路とWntシグナル経路のクロストーク
シュペーマンのオーガナイザーの形成には、TGF-βシグナル経路とWntシグナル経路の相互作用が必要です。これらの経路は、胚の動物極から植物極にかけての細胞の運命を決定します。
オーガナイザー分子ではないが重要な中胚葉マーカー
Brachyury
(マウス) anterior visceral endodermで発現する分子
Otx2
- Sequential roles for Otx2 in visceral endoderm and neuroectoderm for forebrain and midbrain induction and specification M Rhinn 1, A Dierich, W Shawlot, R R Behringer, M Le Meur, S L Ang Development . 1998 Mar;125(5):845-56. doi: 10.1242/dev.125.5.845.
nodal
- Nodal is a novel TGF-beta-like gene expressed in the mouse node during gastrulation X Zhou 1, H Sasaki, L Lowe, B L Hogan, M R Kuehn Nature . 1993 Feb 11;361(6412):543-7. doi: 10.1038/361543a0. 本文有料 要旨より Here we isolate a candidate for the mutated gene which encodes a new member of the transforming growth factor-beta (TGF-beta) superfamily. Expression is first detected in primitive streak-stage embryos at about the time of mesoderm formation. It then becomes highly localized in the node at the anterior of the primitive streak. This region is analogous to chick Hensen’s node and Xenopus dorsal lip (Spemann’s organizer), which can induce secondary body axes when grafted into host embryos (reviewed in refs 5 and 6).
ノーダル遺伝子の同定もネイチャー論文でした。
- Nodal signalling in the epiblast patterns the early mouse embryo J Brennan 1, C C Lu, D P Norris, T A Rodriguez, R S Beddington, E J Robertson Nature . 2001 Jun 21;411(6840):965-9. doi: 10.1038/35082103. 要約より Nodal signals from the epiblast also pattern the visceral endoderm by activating the Smad2-dependent pathway required for specification of anterior identity in overlying epiblast cells.
ノーダルのマウス初期発生における役割の解析もネイチャー論文になりました。
- Nodal signaling from the visceral endoderm is required to maintain Nodal gene expression in the epiblast and drive AVE migration Dev Biol. 2014 Dec 20;400(1):1–9. doi: 10.1016/j.ydbio.2014.12.016 https://pmc.ncbi.nlm.nih.gov/articles/PMC4806383/ conditional mutagenesis of the Nodal gene specifically within the VE leads to reduced Nodal expression levels in the epiblast and incomplete or failed AVE migration
- Nodal Signaling Activates Differentiation Genes During Zebrafish Gastrulation James T Bennett 1, Katherine Joubin 1,3, Simon Cheng 1,2, Pia Aanstad 4,5, Ralf Herwig 4, Matthew Clark 3,4, Hans Lehrach 4, Alexander F Schier 1,6 Dev Biol. 2007 Jan 12;304(2):525–540. doi: 10.1016/j.ydbio.2007.01.012 https://pmc.ncbi.nlm.nih.gov/articles/PMC1885460/
- Nodal specifies embryonic visceral endoderm and sustains pluripotent cells in the epiblast before overt axial patterning Development . 2006 Jul;133(13):2497-505. doi: 10.1242/dev.02413. Epub 2006 May 25. https://pubmed.ncbi.nlm.nih.gov/16728477/
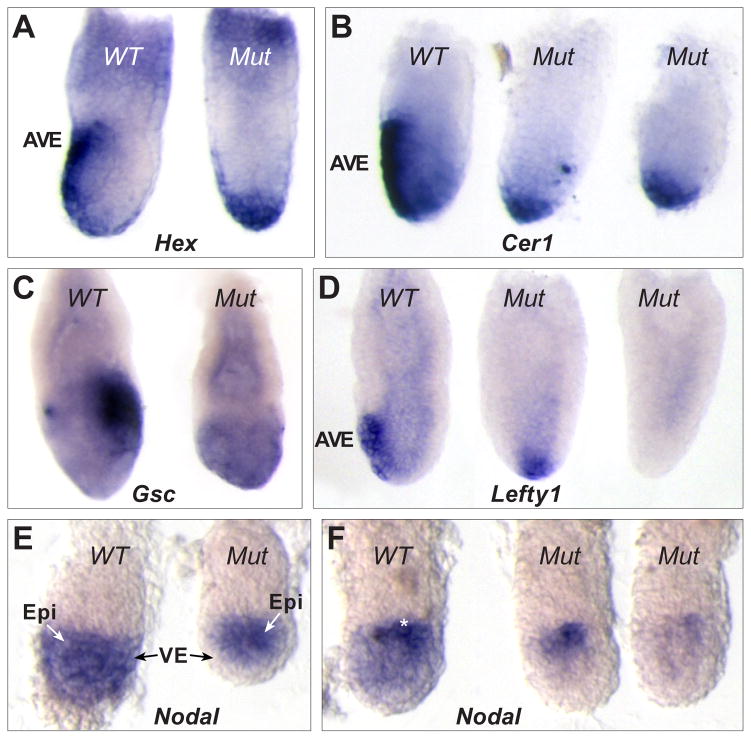
 (B,C) The PrE also expresses the DVE markers Lefty1 (B), Hex and a HexP-GFP reporter transgene(C), whereas Cer1 mRNA is below detectable levels (B). Lefty1 and Hex transcripts are clearly downregulated between E5.0 and 5.25 before they reappear together with Cer1 mRNA in the most distal VE cells at E5.5. https://journals.biologists.com/dev/article/133/13/2497/52434/Nodal-specifies-embryonic-visceral-endoderm-and
(B,C) The PrE also expresses the DVE markers Lefty1 (B), Hex and a HexP-GFP reporter transgene(C), whereas Cer1 mRNA is below detectable levels (B). Lefty1 and Hex transcripts are clearly downregulated between E5.0 and 5.25 before they reappear together with Cer1 mRNA in the most distal VE cells at E5.5. https://journals.biologists.com/dev/article/133/13/2497/52434/Nodal-specifies-embryonic-visceral-endoderm-and - Multiple roles for Nodal in the epiblast of the mouse embryo in the establishment of anterior-posterior patterning Cindy C Lu 1, Elizabeth J Robertson Dev Biol . 2004 Sep 1;273(1):149-59. doi: 10.1016/j.ydbio.2004.06.004. https://pubmed.ncbi.nlm.nih.gov/15302604/
- The Foxh1-dependent autoregulatory enhancer controls the level of Nodal signals in the mouse embryo Dominic P. Norris, Jane Brennan, Elizabeth K. Bikoff, Elizabeth J. Robertson 15 July 2002 https://journals.biologists.com/dev/article/129/14/3455/41736/The-Foxh1-dependent-autoregulatory-enhancer
- nodal expression in the primitive endoderm is required for specification of the anterior axis during mouse gastrulation I Varlet 1, J Collignon, E J Robertson Development . 1997 Mar;124(5):1033-44. doi: 10.1242/dev.124.5.1033.
- A Xenopus nodal-related gene that acts in synergy with noggin to induce complete secondary axis and notochord formation K. D. Lustig, K. Kroll, E. Sun, R. Ramos, H. Elmendorf, M. W. Kirschner Author and article information Development (1996) 122 (10): 3275–3282.
 https://journals.biologists.com/dev/article/122/10/3275/38985/A-Xenopus-nodal-related-gene-that-acts-in-synergy
https://journals.biologists.com/dev/article/122/10/3275/38985/A-Xenopus-nodal-related-gene-that-acts-in-synergy
Cripto
Criptoはアクチビンと結合してアクチビンシグナリングを抑制することはあるようです。しかし、ノーダルに関しては、ノーダルシグナリングの重要な要素コレセプターcoreceptorとしてポジティブに働くようです。ただし、Criptoを介さないノーダルシグナリングもあり得るようです。
- Cripto-independent Nodal signaling promotes positioning of the A-P axis in the early mouse embryo. G. Liguori, A. Borges, A. Borges+9 more · Developmental Biology Volume 315, Issue 2, 15 March 2008, Pages 280-289 2008年3月15日 https://www.sciencedirect.com/science/article/pii/S0012160607016351
その他の論文
- The HPE gene CRIPTO/oep physically interacts with the ligand–receptor complex, and is required for Nodal signal transduction, positioning this protein as a Nodal co-receptor [40–43]. https://en.wikipedia.org/wiki/Nodal_signaling_pathway
- EGF-CFC proteins are membrane bound extracellular factors that serve as essential cofactor in Nodal signaling and in vertebrate development as a whole. This family of cofactors includes One-eyed Pinhead (oep) in Zebrafish, FRL1 in Xenopus, and Cripto and Criptic in mouse and human. https://en.wikipedia.org/wiki/Nodal_signaling_pathway
-
the activity of Nodal requires epidermal growth factor-Cripto/FRL-1/Cryptic (EGF-CFC) co-receptors such as Cripto and Cryptic (Tdgf1 and Cfc1 –Mouse Genome Informatics) (Reissmann et al., 2001; Yan et al.,2002; Yeo and Whitman, 2001). In particular, membrane-boundCripto appears to recruit Nodal to an activin receptor complexcomposed of a dimer of the type I serine-threonine receptor Alk4(Acvr1b) and a dimeric type II activin receptor, either ActRII orActRIIB (Acvr2a or Acvr2b – Mouse Genome Informatics). https://www.researchgate.net/publication/51591179_Regulation_of_extra-embryonic_endoderm_stem_cell_differentiation_by_Nodal_and_Cripto_signaling
Mature Nodal binds to the Activin receptors I and II and the co-receptor Cripto/Criptic and phosphorilates Smad2 /3. https://en.wikipedia.org/wiki/Nodal_signaling_pathway
nodalもアクチビンもTGF-βスーパーファミリーの一員です。アクチビン受容体の種類としては、Nodalに特化した受容体としてはALK7があります。Activinは主にALK4を介してシグナルを伝達します。
- The orphan receptor ALK7 and the Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate development. document Genes & developmentEva Reissmann et al.
TGF-βファミリー分子に対する受容体として,7種類のI型受容体,5種類のII型受容体が存在し,それらすべての受容体は細胞内領域にセリン/トレオニンキナーゼドメインを持つ.ジスルフィド結合により二量体を形成しているTGF-βは,2分子のTGF-β I型受容体と2分子のTGF-β II型受容体からなるヘテロ四量体に結合する(図2).リガンドがTGF-β II型受容体に結合すると,TGF-β II型受容体キナーゼはTGF-β I型受容体の膜貫通領域直下に存在するGS領域と呼ばれるグリシンとセリンに富んだドメインのセリンとトレオニン残基をリン酸化し,I型受容体内で不活性化されていたセリン/トレオニンキナーゼ酵素活性が出現する.TGF-βと同様にアクチビンは,アクチビンI型受容体(ALK4)とアクチビンII型受容体(ActRIIまたはActRIIB)が複合体を形成し,BMPの場合BMPI型受容体(ALK2, 3, 6)とBMP II型受容体(ActRII,ActRIIBまたはBMPRII)が複合体を形成する.活性化したI型受容体キナーゼはR-Smad(receptor-regulated Smad)タンパク質のC末端に存在する2個のセリン残基をリン酸化する.
Journal of Japanese Biochemical Society 89(2): 145-153 (2017) doi:10.14952/SEIKAGAKU.2017.890145 TGF-βシグナルを抑制するTMEPAIファミリー 2017年4月25日 https://seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2017.890145/data/index.html
HEX
HEXはHHEXと呼ばれることもありますが同一の遺伝子です(ChatGPT 4o)。hematopoietically expressed homeobox gene から、HexまたはHhexと呼ばれます。
- Hhex has been found to be an important transcription factor in embryogenesis because Hhex−/− mice display embryonic lethality starting at embryonic day E10.5 to E11.5 due to impaired forebrain, liver, and thyroid development.8–10
- Hhex expression is initially seen in the blood islands of the yolk sac at the same time Flk-1 expression is initiated.9
https://pmc.ncbi.nlm.nih.gov/articles/PMC2938236/
HEXが転写因子として、Cerberus(Wnt, BMP, Nodalのアンタゴニスト)やLefty(TGF-β superfamilyメンバーでNodal signalingを調節)の遺伝子を直接(プロモーターに結合して)制御するのかと思ったのですが、そういうわけではないようです。Cerberusに関しては、HEXによる遺伝子の制御は間接的に起こるようです。Leftyに関しては、遺伝子制御に関する報告がなさそうです(ChatGPT4oの回答)。
- AVE protein expression and visceral endoderm cell behavior during anterior–posterior axis formation in mouse embryos: Asymmetry in OTX2 and DKK1 expression Hideharu Hoshino, Go Shioi, Shinichi Aizawa Developmental Biology Volume 402, Issue 2, 15 June 2015, Pages 175-191 Developmental Biology https://www.sciencedirect.com/science/article/pii/S0012160615001748
- An anterior signalling centre in Xenopus revealed by the homeobox gene XHex C.Michael Jones1 jonesm@icr.ac.uk ∙ Joanne Broadbent2 ∙ Paul Q. Thomas3 ∙ James C. Smith4 ∙ Rosa S.P. Beddington3 Current Biology Volume 9, Issue 17p946-S1-954September 09, 1999 https://www.cell.com/current-biology/fulltext/S0960-9822(99)80421-7
80421-7/asset/739823c5-6901-4ee1-b007-33aa46b7fe6b/main.assets/gr2_lrg.jpg) (f) Animal pole view of stage 10.5 gastrula shows that cerberus expression is detected in a wider domain of the deep endoderm than is XHex (b). 背景より In Xenopus, the putative head inducer cerberus is expressed initially in deep endodermal cells adjacent to the involuting tissues of Spemann’s organiser [7]. Is this tissue the equivalent of the anterior visceral endoderm in the early mouse and rabbit embryo? Here, we have addressed this question by analysing the expression and function of XHex, the Xenopus cognate of murine Hex (also known as Prh) [8,9].
(f) Animal pole view of stage 10.5 gastrula shows that cerberus expression is detected in a wider domain of the deep endoderm than is XHex (b). 背景より In Xenopus, the putative head inducer cerberus is expressed initially in deep endodermal cells adjacent to the involuting tissues of Spemann’s organiser [7]. Is this tissue the equivalent of the anterior visceral endoderm in the early mouse and rabbit embryo? Here, we have addressed this question by analysing the expression and function of XHex, the Xenopus cognate of murine Hex (also known as Prh) [8,9].
DEVからAVEへの細胞移動
- Nodal signaling from the visceral endoderm is required to maintain Nodal gene expression in the epiblast and drive AVE migration Dev Biol. 2014 Dec 20;400(1):1–9. doi: 10.1016/j.ydbio.2014.12.016 https://pmc.ncbi.nlm.nih.gov/articles/PMC4806383/
- Induction and migration of the anterior visceral endoderm is regulated by the extra-embryonic ectoderm Tristan A Rodriguez 1, Shankar Srinivas, Melanie P Clements, James C Smith, Rosa S P Beddington Development . 2005 Jun;132(11):2513-20. doi: 10.1242/dev.01847. Epub 2005 Apr 27.
中胚葉誘導と体軸形成との関係
Mesoderm induction and axis formation are tightly correlated processes in developmental biology, particularly during early embryogenesis. Here’s how they are interconnected:
- Mesoderm Induction: This process begins during gastrulation, where signaling molecules like Nodal, BMP, and Wnt help induce the formation of mesoderm from the epiblast. The mesoderm gives rise to various tissues, including muscles, bones, and the circulatory system.
- Organizer and Axis Formation: The Spemann-Mangold organizer (in amphibians) or the Hensen’s node (in mammals) plays a central role in both mesoderm induction and the formation of the body axis. The organizer secretes BMP antagonists (such as Chordin, Noggin, and Follistatin) that create a dorsal-ventral gradient essential for patterning the mesoderm and establishing the anterior-posterior and dorsal-ventral axes.
- Signaling Pathways:
- Nodal signaling: Initiates mesoderm formation and is essential for the primitive streak formation, which defines the body axis (anterior-posterior).
- BMP inhibition by molecules like Chordin and Noggin, secreted from the organizer, is crucial for dorsalization and axis formation. Without this inhibition, mesoderm would not form correctly, leading to a lack of proper axis formation.
In summary, mesoderm induction and axis formation are closely linked, with key signaling molecules from the organizer region guiding both the formation of mesodermal tissue and the establishment of the body axis during early development.
- ChatGPT 4o