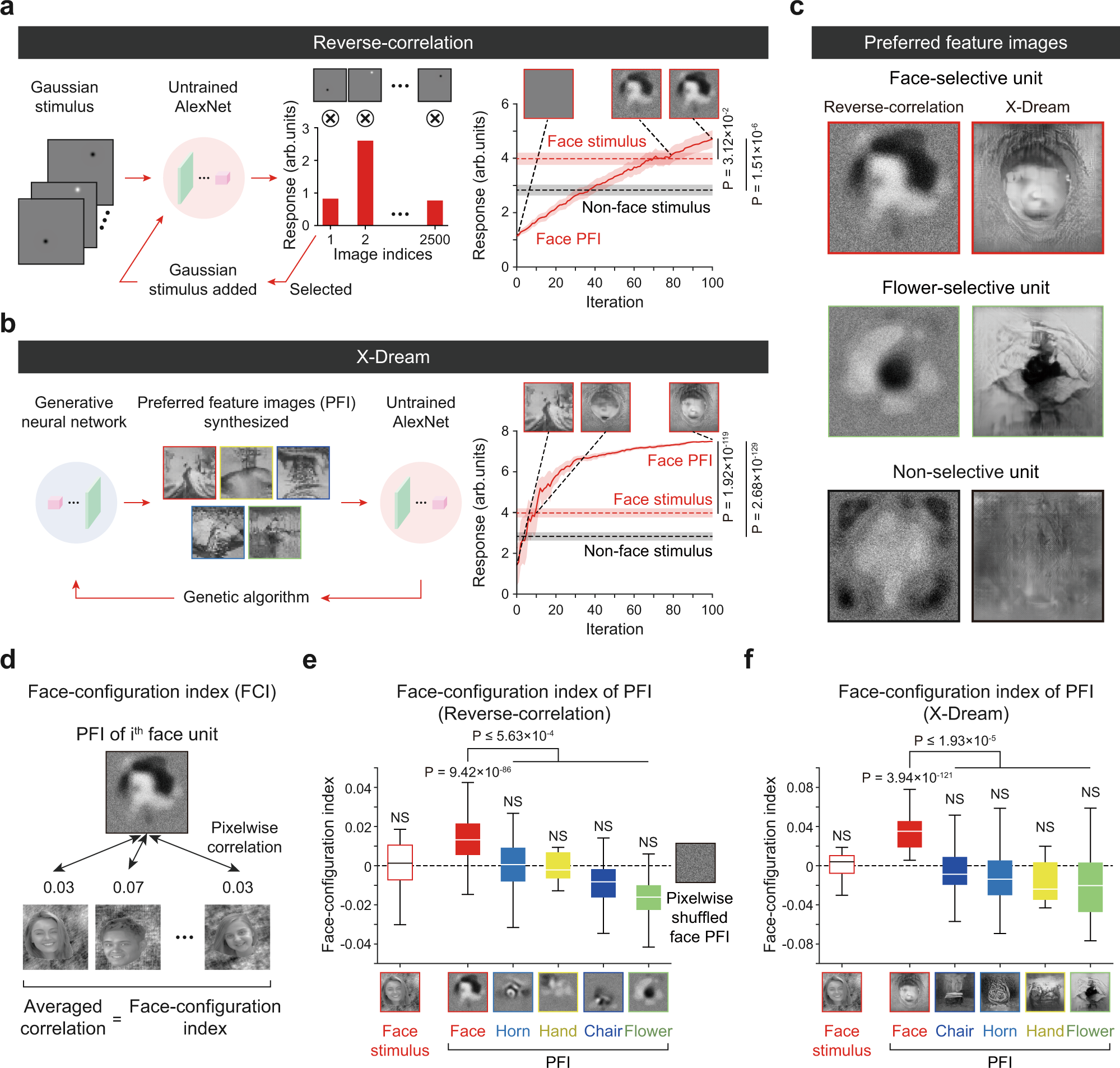
Face detection in untrained deep neural networks Seungdae Baek, Min Song, Jaeson Jang, Gwangsu Kim & Se-Bum Paik Nature Communications volume 12, Article number: 7328 (2021) https://www.nature.com/articles/s41467-021-27606-9 We showed that a biologically inspired DNN develops face-selective units without training, solely from statistical variations of the feedforward projections.
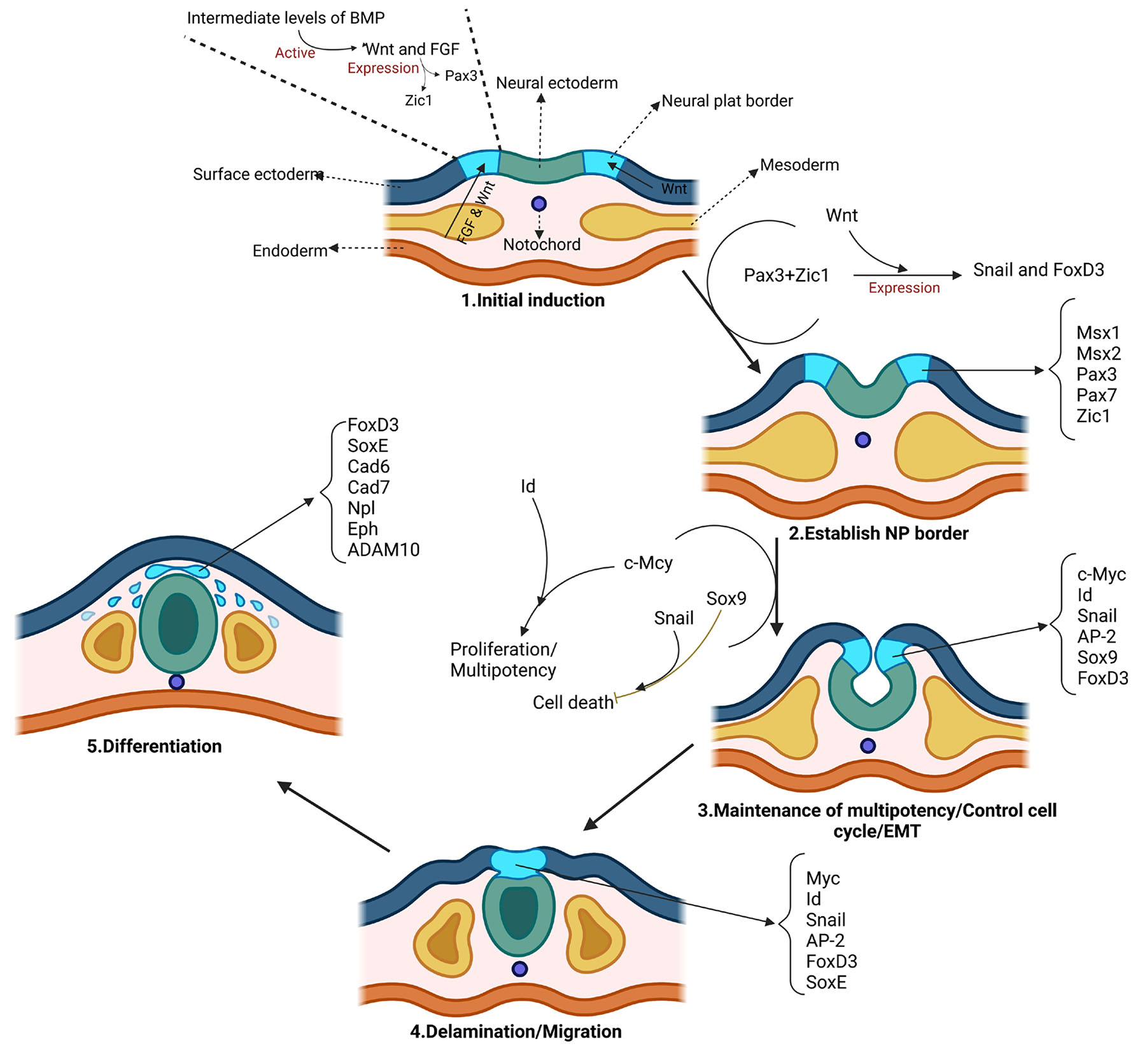
A catenin-dependent balance between N-cadherin and E-cadherin controls neuroectodermal cell fate choices Mechanisms of Development Volume 152, August 2018, Pages 44-56 Mechanisms of Development https://www.sciencedirect.com/science/article/pii/S0925477318300819
N- and E-cadherins in Xenopus are specifically required in the neural and non-neural ectoderm, respectively, for F-actin assembly and morphogenetic movements Development (2009) 136 (8): 1327–1338. https://doi.org/10.1242/dev.031203 15 April 2009
Fig. 4. Shh- and Noggin-expressing cells prevent or down-regulate Slug expression in the dorsal neural tube and influence the expression of other dorsal neural tube genes.
Figure 2 Induction of Floor Plate Cells in Response to Shh and Chordin In Vivo (N and O) Coexpression of (N) Shh and (O) chordin in the HH st-10 caudal notochord.
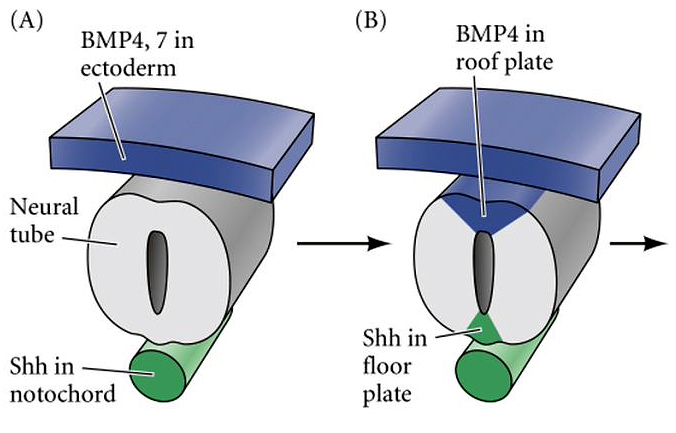
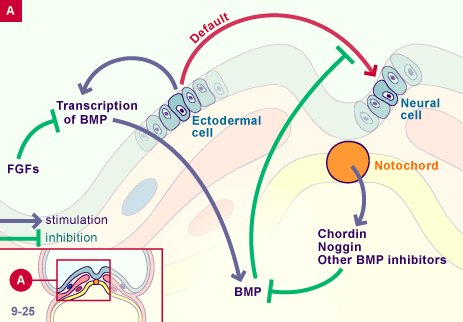
Effects of Shh and Noggin on neural crest formation demonstrate that BMP is required in the neural tube but not ectoderm Development (1998) 125 (24): 4919–4930.
BMP inhibition initiates neural induction via FGF signaling and Zic genes Leslie Marchal, Guillaume Luxardi, Virginie Thomé, and Laurent Kodjabachian kodja@ibdml.univ-mrs.frAuthors Info & Affiliations Edited by Igor B. Dawid, National Institute of Child Health and Human Development, Bethesda, MD, and approved August 19, 2009 October 13, 2009 106 (41) 17437-17442 https://www.pnas.org/doi/10.1073/pnas.0906352106 neural induction is controlled by BMP inhibition, which activates directly, and, via FGF instructive activity, early neural regulators such as Zic genes.
The response of early neural genes to FGF signaling or inhibition of BMP indicate the absence of a conserved neural induction module Crystal D Rogers 1, George S Ferzli 1, Elena S Casey BMC Dev Biol. 2011 Dec 15;11:74. doi: 10.1186/1471-213X-11-74
Combinatorial Fgf and Bmp signalling patterns the gastrula ectoderm into prospective neural and epidermal domains Development. 2004 Aug;131(15):3581–3592. doi: 10.1242/dev.01227 https://pmc.ncbi.nlm.nih.gov/articles/PMC2789263/
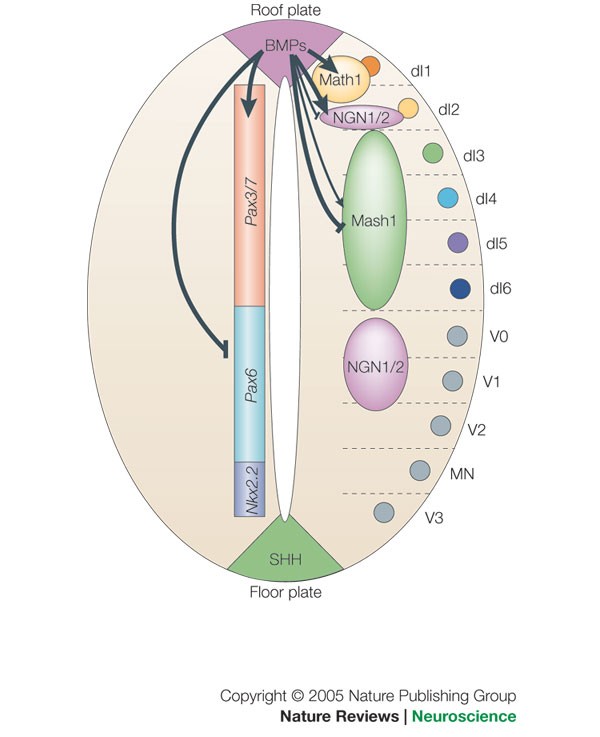
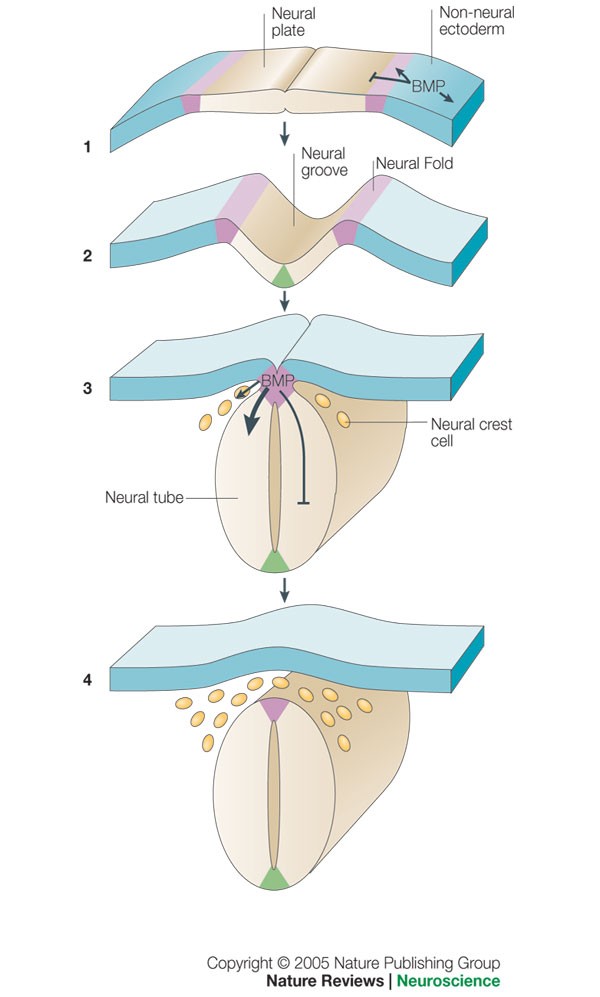
Bone morphogenetic protein signalling and vertebrate nervous system development 01 December 2005 Aimin Liu & Lee A. Niswander Nature Reviews Neuroscience volume 6, pages945–954 (2005) https://www.nature.com/articles/nrn1805
Neural Plate Patterning by Secreted Signals Oliver Wessely ∙ E.M De Robertis NEURON Volume 33, Issue 4p489-491February 14, 2002 derobert@hhmi.ucla.edu https://www.cell.com/fulltext/S0896-6273%2802%2900596-2
22.2 Early embryonic development of the nervous system: Formation of the neural tube and neural crest https://embryology.ch/en/organogenesis/nervous-system/early-embryonic-development-of-the-nervous-system/molecular-mechanisms-in-the-early-development-of-cns/dorso-ventral-polarity-of-neural-tube.html?p=2.2
Current Biology Volume 12, Issue 1p47-52January 08, 2002 Opponent Activities of Shh and BMP Signaling during Floor Plate Induction In Vivo https://www.cell.com/current-biology/fulltext/S0960-9822(01)00631-5
Noggin-mediated antagonism of BMP signaling is required for growth and patterning of the neural tube and somite Jill A. McMahon1, Shinji Takada1,4, Lyle B. Zimmerman3,5, Chen-Ming Fan2, Richard M. Harland3, and Andrew P. McMahon1,6 doi: 10.1101/gad.12.10.1438 Genes & Dev. 1998. 12: 1438-1452
A Mechanism Regulating the Onset of Sox2 Expression in the Embryonic Neural Plate 神経板の最も初期の決定的なマーカーは転写因子 Sox2 であり、これは ERNI、ジェミニン、BERT、HP1α、HP1β、およびクロマチンリモデリング酵素 Brm 間の相互作用によって制御されます。 PLoS Biology C. Papanayotou et al. 171 Citations 2008
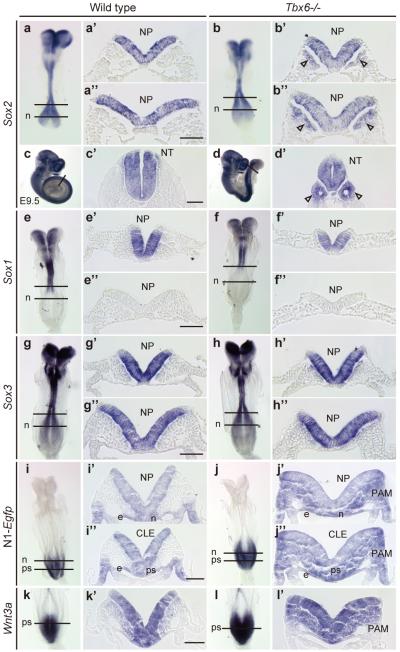
Nature. 2011 Feb 17;470(7334):394–398. doi: 10.1038/nature09729 Tbx6-dependent Sox2 regulation determines neural vs mesodermal fate in axial stem cells
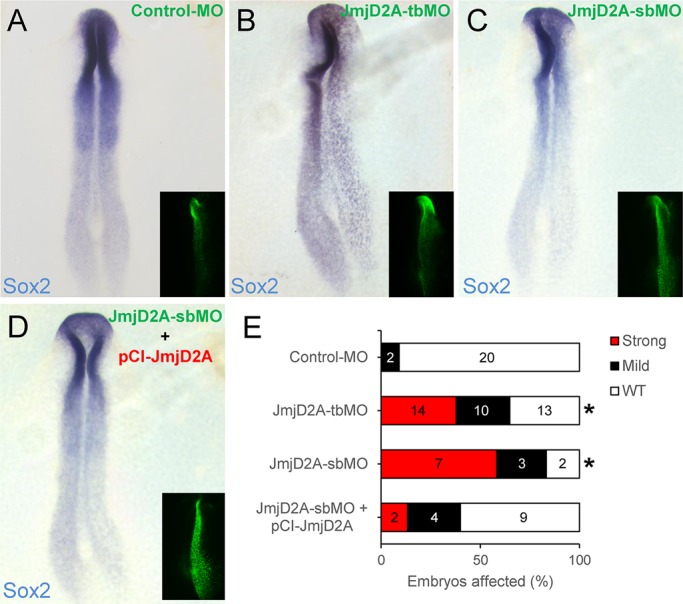
Epigenetic activation of Sox2 gene in the developing vertebrate neural plate Mol Biol Cell. 2016 Jun 15;27(12):1921–1927. doi: 10.1091/mbc.E16-01-0042 https://pmc.ncbi.nlm.nih.gov/articles/PMC4907725/
A role for SOX1 in neural determination. SRY に関連する HMG ボックス タンパク質である SOX1 は、神経運命が決定された外胚葉細胞で発現される最も初期の転写因子の 1 つであり、胚の中枢神経系 (CNS) の分裂中の神経前駆細胞を定義します。 Development L. Pevny et al. 496 Citations 1998 https://journals.biologists.com/dev/article/125/10/1967/39814/A-role-for-SOX1-in-neural-determination 図1 Immunohistochemical analysis of SOX1 during mouse and rat embryogenesis. (A) Transverse section through the anterior region of a 7.5 dpc mouse embryo. SOX1 labeling is restricted to the neural plate. (B) Transverse section through the anterior region of a 2 somite mouse embryo. SOX1 labeling is restricted to the neural folds. (C) Transverse section through the posterior region of a 10-12 somite mouse embryo. SOX1 labeling is restricted to the neural tube. (D) Transverse section through the thoracic region of a 20 somite mouse embryo. SOX1 labeling is detected in the early neural tube but is excluded from the ventral midline region of the floor plate (FP, arrow).Sox2 図1aをみるとSOX1が神経板に局在している様子がハッキリとわかります。神経管が形成された時期の図1cでは神経管に特異的に発現しています。
Inhibition of BMP signaling is sufficient to induce neural markers, including SOX1, in ectodermal explants, suggesting that neural fate is the default state in the ectoderm when BMP signaling is blocked . The use of BMP inhibitors like Noggin and DMH1 can effectively induce SOX1 expression in human-induced pluripotent stem cells (hiPSCs), highlighting the role of BMP antagonists in neural induction .
FGF signaling is necessary for maintaining the early domain of sox gene expression, including SOX1, in neural tissue. Without FGF signaling, this domain can revert to non-neural fates. The combined inhibition of BMP and TGF-β pathways, using small molecule inhibitors, allows for precise regulation of SOX1 expression, which is crucial for proper neural induction and lineage specification.
XASH-3
XASH-3は、神経誘導の非常に早い段階で発現し、神経板の位置を示すマーカーとして機能します。
XASH-3, a novel Xenopus achaete-scute homolog, provides an early marker of planar neural induction and position along the mediolateral axis of the neural plate. Development K. Zimmerman et al. 134 Citations 1993 https://journals.biologists.com/dev/article/119/1/221/37847/XASH-3-a-novel-Xenopus-achaete-scute-homolog XASH-3 は、アフリカツメガエルの神経板の内外軸に沿った神経誘導と位置の初期マーカーです。 Comparison of XASH3 and N-CAM expression in late-gastrula stage embryos. XASH3 (A,B,C) and N-CAM (D,E) expression in stage 14 embryos was determined by whole-mount in situ hybridization (A,D). Cross-sections of similarly staged embryos are also shown (B,C and E). Open arrow in A indicates XASH3 expression lateral to the neural plate; closed arrow indicates the mediolaterally restricted stripe revealed by crosssection in B, and the arrowhead indicates the transverse ‘eyebrow’ stripe revealed by cross-section in C. In A, anterior is towards 11 o’clock and, in D, it is towards 1 o’clock. Note the restricted expression of XASH3 along the mediolateral axis of the neural plate (B) in comparison to that of N-CAM (E).
Gbx2
Gbx2は、神経板の後方化に関与し、神経板のパターン形成において重要な役割を果たします。
The posteriorizing gene Gbx2 is a direct target of Wnt signalling and the earliest factor in neural crest induction Gbx2 は神経堤誘導における最も初期の因子であり、神経板の後部化に役割を果たします。 Development Bo Li et al. 146 Citations 2009 Info In Vitro Trial Info Highly Cited https://journals.biologists.com/dev/article/136/19/3267/65330/The-posteriorizing-gene-Gbx2-is-a-direct-target-of Gbx2 is expressed in posterior ectoderm that includes the prospective neural crest. (A-C) Hypothesis of neural crest (NC)induction by the posteriorizing activity of Gbx2. (D-S) In situ hybridization at the indicated stages for the indicated genes. (D-G)Dorsal view, anterior to the top. (H-K) Transverse sections. (L-O) Lateral view, anterior to the left. (P-R) Dorsal view, anterior to the top. Arrowhead,NC; arrow, gap in Gbx2 expression. (S) Detail of the neural fold region in a lateral view, anterior to the top, midline to the right.(T) Summary of Gbx2 and Snail2 expression at stage 16. Anterior to the top, midline to the right. Different tones of purple denote different levels of Gbx2 expression. Blue, NC.
N-CAM
N-CAM RNAの発現は、神経誘導の初期応答として、神経板に局在します。
Expression of Xenopus N-CAM RNA in ectoderm is an early response to neural induction. N-CAM RNA の発現は、胚葉形成中に神経板に局在し、外胚葉における神経の関与のマーカーとなります。 Development C. Kintner et al. 468 Citations 1987 https://journals.biologists.com/dev/article/99/3/311/51817/Expression-of-Xenopus-N-CAM-RNA-in-ectoderm-is-an
pou5f3/pou2
Involvement of an Oct4-related PouV gene, pou5f3/pou2, in neurogenesis in the early neural plate of zebrafish embryos. Pou5f3 は、ゼブラフィッシュの胚において、前神経クラスターにおける初期の神経発生を促進し、後期の神経発生を負に制御します。 Developmental biology Chihiro Inomata et al. 6 Citations 2019
Xsnail
Relationship between gene expression domains of Xsnail, Xslug, and Xtwist and cell movement in the prospective neural crest of Xenopus. Xsnail は、神経誘導における神経板の決定のための最も初期のマーカーです。 Developmental biology C. Linker et al. 103 Citations 2000
neurogenin
The activity of neurogenin1 is controlled by local cues in the zebrafish embryo. ゼブラフィッシュ胚の初期神経板におけるニューロジェニン1の発現はDelta/Notchシグナル伝達によって調節され、異所性ニューロンの発達につながる可能性があります。 Development P. Blader et al. 259 Citations 1997
Multiple regulatory elements with spatially and temporally distinct activities control neurogenin1 expression in primary neurons of the zebrafish embryo ゼブラフィッシュの胚におけるニューロジェニン1の発現は複数の調節要素によって制御され、特定のニューロン前駆細胞における発現を促進し、神経板におけるその発現を調節します。 Mechanisms of Development P. Blader et al. 164 Citations 2003
A novel zebrafish bHLH gene, neurogenin3, is expressed in the hypothalamus. ニューロジェニンは決定因子として作用し、脊椎動物の神経発生における分化遺伝子の発現を開始します。 Gene Xukun Wang et al. 43 Citations 2001
その他
A gene regulatory network for neural induction 神経誘導のための遺伝子制御ネットワークは、175 個の転写制御因子とそれらの間の 5,614 個の予測される相互作用で構成され、信号への最初の露出から成熟した神経板マーカーの発現までの細かい時間的ダイナミクスを備えています。 eLife K. Trevers et al. 5 Citations 2021
The Netrin receptor Neogenin is required for neural tube formation and somitogenesis in zebrafish. ネオジェニンは、ゼブラフィッシュの胚における神経管形成と体節形成において重要な役割を果たし、神経外胚葉細胞と中胚葉細胞の両方の細胞極性または移動方向性を決定します。 Developmental biology D. Mawdsley et al. 65 Citations 2004







00631-5/asset/4b40e407-6a99-470e-9479-f37c3eb71332/main.assets/gr2_lrg.jpg)











00596-2/asset/21dcc826-cd55-4248-a17e-968b247791b3/main.assets/gr1_lrg.jpg)

00631-5/asset/3329ee04-8b3c-4804-8945-543f4ce17880/main.assets/gr2.jpg)